- Infectious Diseases of Livestock
- Part 1
- African animal trypanosomoses
- Vectors: Ticks
- Vectors: Tsetse flies
- Vectors: Muscidae
- Vectors: Tabanidae
- Vectors: Culicoides spp.
- Vectors: Mosquitoes
- Classification, epidemiology and control of arthropod-borne viruses
- Special factors affecting the control of livestock diseases in sub-Saharan Africa
- The control of infectious diseases of livestock: Making appropriate decisions in different epidemiological and socioeconomic conditions
- Infectious diseases of animals in sub-Saharan Africa: The wildlife⁄livestock interface
- Vaccination: An approach to the control of infectious diseases
- African animal trypanosomoses
- Dourine
- Trichomonosis
- Amoebic infections
- GENERAL INTRODUCTION: COCCIDIA
- Coccidiosis
- Cryptosporidiosis
- Toxoplasmosis
- Besnoitiosis
- Sarcocystosis
- Balantidiosis
- Leishmaniosis
- Neosporosis
- Equine protozoal myeloencephalitis
- GENERAL INTRODUCTION: BABESIOSES
- Bovine babesiosis
- Equine piroplasmosis
- Porcine babesiosis
- Ovine babesiosis
- GENERAL INTRODUCTION: THEILERIOSES OF CATTLE
- East Coast fever
- Corridor disease
- Zimbabwe theileriosis
- Turning sickness
- Theileria taurotragi infection
- Theileria mutans infection
- Theileria annulata theileriosis
- Theileriosis of sheep and goats
- Theileria buffeli⁄orientalis infection
- Non-pathogenic Theileria species in cattle
- GENERAL INTRODUCTION: RICKETTSIAL, CHLAMYDIAL AND HAEMOTROPIC MYCOPLASMAL DISEASES
- Heartwater
- Lesser known rickettsial infections in animals and humans
- Chlamydiosis
- Q fever
- Eperythrozoonosis
- Bovine Haemobartonellosis
- Potomac horse fever
- GENERAL INTRODUCTION: ANAPLASMOSES
- Bovine anaplasmosis
- Ovine and caprine anaplasmosis
African animal trypanosomoses
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here

African animal trypanosomoses
R J CONNOR AND P VAN DEN BOSSCHE
Introduction
The trypanosomoses are diseases of humans and domestic animals that result from infection with parasitic protozoa of the genus Trypanosoma. Trypanosomes parasitize all classes of vertebrates: fish, amphibians, reptiles, birds and mammals. The parasites, with the exception of Trypanosoma equiperdum, the cause of dourine, are transmitted from host to host by haematophagous vectors, and usually cause little appreciable harm to either the vector or the vertebrate host. However, several species of trypanosomes which parasitize mammals are less well adapted and commonly cause disease.
The trypanosomoses form a group of diseases, each species of pathogenic trypanosome causing the disease trypanosomosis. The course of a trypanosomal infection varies considerably and depends upon both the species of trypanosome and the host involved. Trypanosomosis is generally characterized by the intermittent presence of parasites in the blood and intermittent fever. Anaemia usually develops in affected animals, and this is followed by loss of body condition, reduced productivity and, often, high mortality.
The first report that associated trypanosomes with disease was made from India by Evans81 in 1880. He found trypanosomes in the blood of camels and horses which were affected by a disease known locally as ‘surra’, and which is now known to be caused by Trypanosoma evansi. Subsequently, Bruce35 made the major discovery in Zululand, South Africa, that trypanosomes were the causal organisms of ‘nagana’, or tsetse fly disease. The early review of trypanosomes and trypanosomoses by Laveran and Mesnil,155 translated from the French by Nabarro in 1907, was followed by Wenyon’s316 contribution in 1926. Subsequent major reviews of trypanosomes and African trypanosomoses were made by Mulligan and Potts,204 Ford,85 Hoare,111 Jordan,136 and Stephen.274 The considerable interest in trypanosomoses arises from their importance and the intriguing biology of the causal organisms.
Two forms of human trypanosomosis exist: Chagas’ disease occurs in Central and South America and is transmitted by blood-sucking reduviid bugs, certain small wild animals and dogs harbouring the infection. The second form is human sleeping sickness. This occurs in Africa and is transmitted by blood-sucking flies of the genus Glossina, commonly known as ‘tsetse flies’ or simply as ‘tsetse’. The majority of animal diseases caused by trypanosomes occur in the tropics. In Africa, several species of tsetse-transmitted trypanosomes cause African trypanosomoses in domestic animals, which in southern Africa are collectively known as ‘nagana’, a word derived from the Zulu ‘nakane’ meaning tsetse fly disease. An important form of trypanosomosis known as dourine, caused by T. equiperdum, also occurs in southern Africa, but has no arthropod vector (see Dourine). ‘Surra’ is transmitted by biting flies other than tsetse flies and, although it occurs in many parts of the tropics, including northern Africa, it is not present in southern Africa.
Tsetse flies have recently been discovered in Saudi Arabia, 77 but they are only known to be of importance in Africa, south of the Sahara, where the diseases they transmit are responsible for great economic loss (see Vectors: Tsetse flies).
The large populations of wild animals, which have thrived for millennia in the tsetse-infested tracts of Africa, have evolved with these flies and the trypanosomes they transmit. Hosts and parasites have become mutually adapted and coexist in a balanced relationship. Humans first brought domestic animals into the tsetse fly belts of Africa relatively recently, and although humpless cattle of the Bos taurus type, from which the present-day taurine breeds such as the West African Shorthorn, N’Dama, Muturu and Baoule are believed to have descended, were introduced into northern and western Africa from 4 500 BC onwards, the humped Zebu, Bos indicus type, arrived some 3 000 years later, and did not reach central and southern Africa until around AD 700.80 Goats and sheep were introduced at about the same time. Because of this relatively recent introduction, the relationship between tsetse-transmitted trypanosomes and domestic animals has not fully evolved and infection with these parasites frequently produces disease. However, the West African humpless cattle have had longer to adapt than Zebu cattle and this may explain why they possess the trait of trypanotolerance. They are able to live without drug treatment in tsetse-infested areas where other cattle die. Furthermore, some indigenous breeds of goats and sheep, such as the Dwarf goats and Djallonké sheep of West Africa and the small East African breeds, are more trypanotolerant than are exotic breeds.211
The ravages of nagana have long been recognized by the inhabitants of southern Africa, and early attempts to introduce livestock into tsetse-infested areas were unsuccessful as draught animals and other stock succumbed to the disease. The devastation which resulted from the rinderpest pandemic of the 1890s (see Rinderpest) destroyed almost entire populations of wild animals and millions of cattle. Without hosts on which to feed, tsetse disappeared from large areas. With the constraint of tsetsetransmitted trypanosomosis removed, settlement in Zimbabwe was rapid. Some 25 000 cattle had survived the rinderpest catastrophy, and after restocking there were over 2,3 million cattle in the country by 1929.41 However, at about this time tsetse were dispersing from residual pockets, and trypanosomosis again became a problem for livestock owners. By 1931, tsetse were spreading at a rate of 2 500 km2 annually, 42 and game elimination to control tsetse began in 1932.43 Since then, strenuous efforts have been made to contain the tsetse fly (see Vectors: Tsetse flies). In many other parts of southern Africa, livestock owners have also had to live with the tsetse fly and its consequences.
Tsetse infest 10 million square kilometres and affect 37 countries, which makes African animal trypanosomosis a problem of truly continental magnitude (Figure 12.1). They live in frost-free areas that have an annual rainfall of 650mm or more. In arid, marginal habitats, tsetse only exist in the better wooded and better watered strips where the host species concentrate during critical times, such as in the late, hot, dry season. In areas of mixed agriculture, cattle are valued for the milk, draught power and the manure which they provide. They also represent a means of investment. In mostrural areas people rely on poultry and small ruminants for meat, cattle only being slaughtered for important social ceremonies or to realize large sums of money. Most of the settled areas of the tsetse fly belts of southern Africa are used for traditional mixed farming, but the presence of tsetse seriously handicaps development.177 The general improvement of agricultural production depends on the greater use of animal traction, in place of manual tillage, enabling larger areas to be more efficiently cultivated. However, animals alone cannot ensure this development. The availability of credit schemes and supplies of seed, fertilizer and implements are also needed before the potential can be realized. The presence of tsetse is, nevertheless, the largest single obstacle to progress in affected areas.
The distribution of tsetse-transmitted bovine trypanosomosis in southern Africa is largely determined by the distribution of tsetse in three major fly belts. The first fly belt, with Glossina morsitans morsitans and Glossina pallidipes as the main tsetse species, links tsetse foci in Malawi with the large fly belt common to eastern Zambia, Zimbabwe and Mozambique.
The Glossina morsitans centralis fly belt covers western Zambia, parts of western Angola, the Kwando River drainage in Namibia’s Eastern Caprivi district and the Okavango Delta in Botswana. Finally, a third fly belt with the species Glossina austeni and Glossina brevipalpis covers parts of Mozambique’s southernmost Matutuine district and north-eastern KwaZulu-Natal in South Africa.
Concerted efforts to control tsetse over the past 50 years have resulted in significant changes in the distribution of tsetse and tsetse-transmitted trypanosomosis. Unfortunately, few of these achievements have been sustained. In many countries of southern Africa, the current distribution of tsetse and, hence, tsetse-transmitted trypanosomosis is not much different from the ecological limits of the fly distribution.
The epidemiology of bovine trypanosomosis differs significantly between countries. Generally speaking, three different types of epidemiological situations can be distinguished. First, in Namibia and Zimbabwe, progressive tsetse control operations have gradually pushed the tsetse front to its current position. The interface between tsetse and cattle occurs at the edge of the artificial barrier, preventing tsetse from reinvading cleared areas, with the possibility of cattle entering tsetse-infested country. Second, in Botswana, Malawi and South Africa, interaction between tsetse and cattle occurs at the edge of the ecological limits of tsetse distribution. In principle, cattle are not able to enter tsetse-infested land. Finally, in Angola, Mozambique and Zambia, cattle live in tsetse-infested areas. They are surrounded by tsetse flies and are subjected to continuous challenge.
In Namibia, bovine trypanosomosis is most prevalent along the Kwando River on the western boundary of the Eastern Caprivi.295 The deployment of odour-baited, insecticide- treated targets along the northern part of the Kwando River has resulted in a significant decline in the incidence of trypanosomosis. Occasionally, cases of nagana are detected in cattle grazing in the vicinity of Katima Mulilo, situated near the border with Zambia. These latter infections are assumed to originate from the neighbouring Sesheke area of Zambia: cattle that graze along the Zambezi River are probably challenged by tsetse that cross the river.
In Zimbabwe, tsetse has been controlled successfully and control efforts have been sustained. This is partly due to the country’s favourable position at the edge of the natural limits of the tsetse distribution. Since 1981, approximately 55 000 km2 of tsetse-infested land has been cleared of tsetse using a variety of methods.268 A combination of odourbaited targets and insecticide-treated cattle constitute ‘barrier’ to reinvasion, confining tsetse to areas less suitable for agriculture or along international borders, and preventing them from invading previously cleared areas.
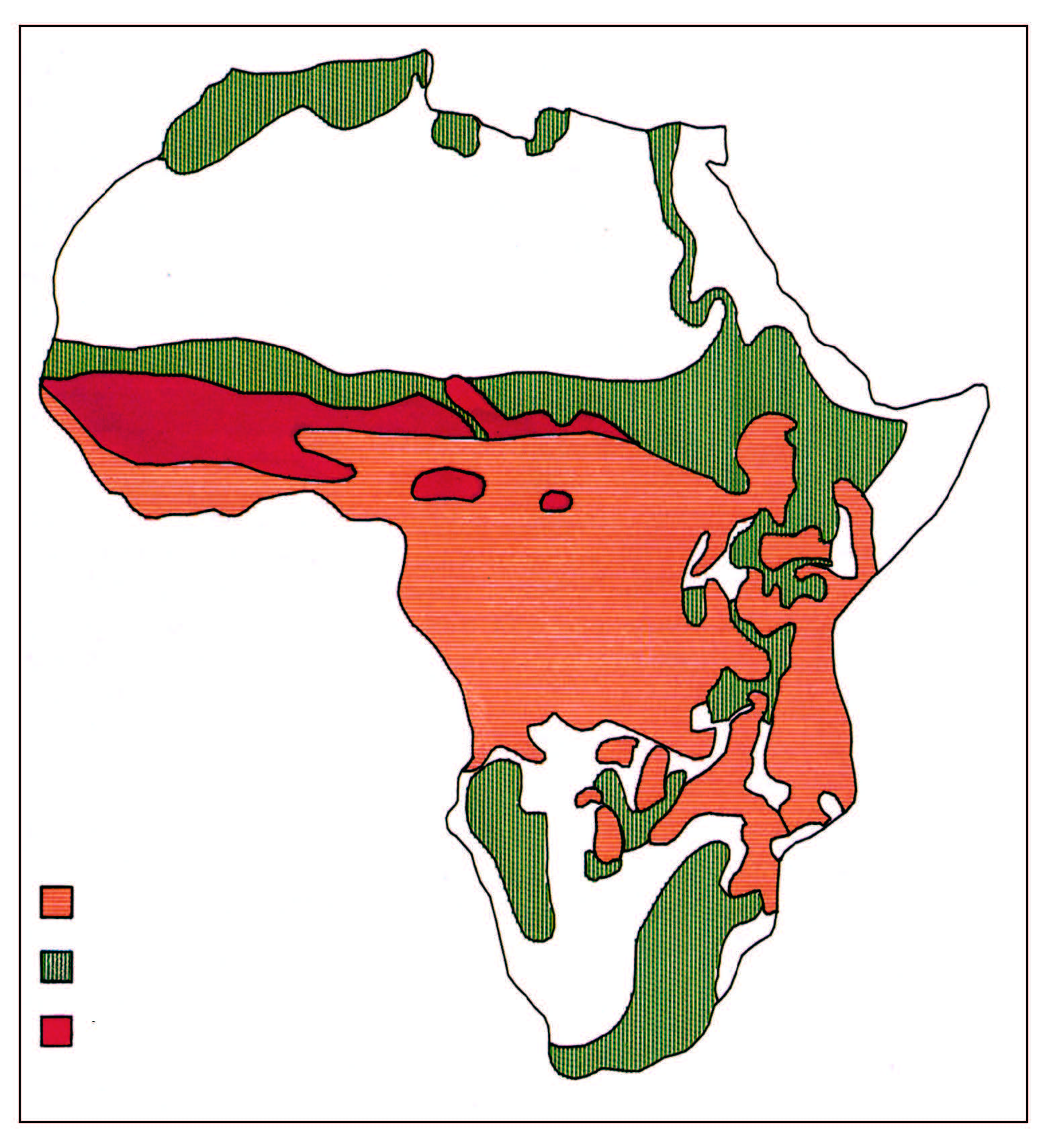
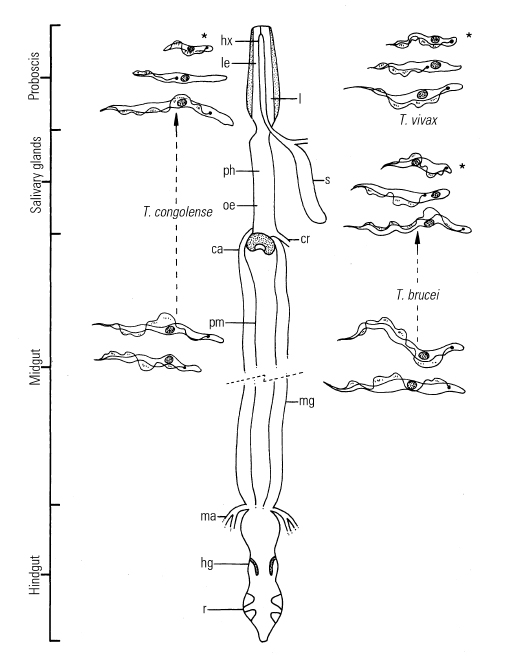
Figure 12.2 Life cycles of African pathogenic trypanosomes
ca = cardia (= proventriculus)
cr = crop duct
hg = hindgut
hx = hypopharynx
l = labium
le = labrum-epipharynx
hx + l + le = proboscis
ma = Malpighian tubes
mg = midgut
oe = oesophagus
ph = pharynx
pm = peritrophic membrane
r = rectum
s = salivary glands
* = infective metatrypanosomes
By the late 1990s, bovine trypanosomosis was prevalent only in the northern and eastern parts of the country. In these areas, cattle contract the disease as they graze within or along the edge of the barrier294 where they are exposed to tsetse fly. Challenge is generally low and bovine trypanosomosis does not constitute a major constraint to animal health and production. Notwithstanding the favourable disease situation, the trypanosomosis threat is real. A breakdown of the barrier to reinvasion could expose over one million highly susceptible cattle in the commercial and communal farming sectors to the risk of trypanosomosis. Continuous disease surveillance is essential. The speed with which tsetse fly are capable of reinvading cleared areas was demonstrated on an experimental basis when a portion of the target barrier was removed in the northeast of the country.304 The impact of the breakdown of control on animal health was also clearly shown.
Tsetse flies infest two areas in Botswana, the Kwando– Linyanti–Chobe fly belt and the Okavango fly belt. Most of the tsetse-infested area comprises wildlife zones that are free of cattle. Because cattle–fly contact is low, bovine trypanosomosis is not a major disease in Botswana. The more immediate risk is to the large number of tourists who visit the wildlife areas, as they face the risk of contracting sleeping sickness.
In Malawi, where the human population density is high, the pressure on land and the concomitant clearing of vegetation for cultivation, confines tsetse to game reserves, national parks or forest reserves. Consequently, cases of nagana are restricted to areas surrounding the tsetseinfested zones and areas adjacent to neighbouring Zambia or Mozambique where tsetse is present.292 The prevalence of infection depends upon the cattle–fly contact and varies between areas and seasons. It decreases with increasing distance from the tsetse infestation. Although bovine trypanosomosis can assume epidemic proportions, it is localized and does not constitute a major threat to livestock production in Malawi. The ever-increasing demand for land will probably further reduce suitable tsetse habitat. However, the expansion of the human population will compel people to live at the edge of tsetse infestations, which is likely to result in an increased incidence of nagana.
In the late 1850s, tsetse flies infested the Limpopo, Mpumalanga and KwaZulu-Natal provinces of South Africa, 110 but the rinderpest pandemic led to the complete disappearance of tsetse from these areas. However, in parts of Zululand, including the present-day game reserves of north-eastern KwaZulu-Natal, pockets of wild animals and tsetse survived, and by 1905 nagana was again threatening livestock in the region. Glossina pallidipes disperse into farming areas causing serious epidemics. The advent of organochlorine insecticides provided a suitable means to remove tsetse, and between 1945 and 1952 G. pallidipes was eradicated from Zululand by large-scale aerial spraying operations.65 Thereafter, the two species found in Zululand, G. austeni and G. brevipalpis, were responsible for sporadic cases of bovine trypanosomosis.142 Despite the restricted distribution of tsetse, a widespread outbreak of bovine trypanosomosis occurred in 1990.13
In Angola, nagana is a serious constraint. Over a quarter of the country is tsetse-infested95 and the 3,5 million cattle are kept mainly in the south-western, tsetse-free areas, although there is a small population of trypanotolerant cattle in Cabinda Province. Control measures are currently limited to the use of trypanocidal drugs.
Tsetse fly are a serious threat to livestock development in about two-thirds of Mozambique. More than two decades of civil war reduced the cattle population by more than 80 per cent and made bovine trypanosomosis less prevalent than before. Currently, most of Mozambique’s livestock is distributed in areas where tsetse is either absent or present at low levels. Nevertheless, bovine trypanosomosis constitutes a threat to the development of rural areas and could be a serious constraint to restocking programmes.
In Zambia, on the other hand, bovine trypanosomosis is a disease of national importance. Three-fifths of the country provide suitable tsetse habitat. Despite its long history of tsetse control, large-scale operations have met with limited long-term success. This is largely attributable to the absence of natural barriers, which results in a continuous threat of reinvasion of cleared areas by tsetse from adjacent infested zones. In contrast to most other countries in southern Africa, but in common with countries in eastern Africa, cattle are kept in tsetse-infested areas. Under these conditions, disease challenge and prevalence of infection are generally high. Consequently, bovine trypanosomosis constitutes a major constraint to rural development in large areas of western, southern and eastern Zambia.44
Early work on trypanosomosis, much of it conducted in southern Africa, concentrated on describing the trypanosomes and studying the natural history of the parasites, their vectors and their hosts. The greatest advances in our knowledge of trypanosomosis over the past two decades have been made in the areas of molecular biology and immunology, rather than at the field level. Although extremely valuable research has been conducted in carefully controlled laboratory experiments, it is often difficult to relate the results to the field situation.
Aetiology and life cycle
Trypanosomes are protozoan parasites of the genus Trypanosoma, order Kinetoplastida, and have, as characteristic organelles, a kinetoplast and a flagellum. Typically, trypanosomes are digenetic parasites and thus require two hosts to complete their life cycle: they multiply in the blood, tissues or body fluids of a vertebrate host and, with the exception of T. equiperdum which is venereally transmitted, are ingested by a haematophagous invertebrate vector. With a few notable exceptions, a cycle of development and maturation occurs in the vector, after which the parasites are transmitted to another vertebrate host as the vector feeds. Transmission is either by inoculation of trypanosomes with saliva or by contamination of mucosa or broken skin with trypanosomes in the vector’s faecal material, voided during the blood meal. The type of development cycle within the vector determines whether or not infective, metacyclic parasites are present in saliva or faeces. On this basis mammalian trypanosomes are classified into the two broad sections of ‘salivaria’ and ‘stercoraria’.111
In Africa, the pathogenic trypanosomes that cause sleeping sickness in humans and nagana in domestic animals are salivarian, and cyclical development occurs in tsetse flies (Figure 12.2). Transmission of any trypanosome species can take place mechanically without cyclical changes occurring in the vector. In nature, this is effected by biting flies, such as Tabanus, Stomoxys and Lyperosia spp., which feed on more than one animal before repletion. The fly remains infective for only a short time, although in South America and Mauritius (see below) mechanical transmission of Trypanosoma vivax by haematophagous flies other than tsetse has enabled the parasite to become established. Experimentally, trypanosomes may be transmitted by ‘syringe passage’ of infective blood.270 ‘Surra’ is a disease that affects a wide range of host animals, and it occurs in North Africa, the Near and Far East, Central and South America, the Philippines and Mauritius. It is caused by T. evansi, a dyskinetoplastic form of which — known as Trypanosoma equinum — also causes disease in equids in Central and South America, where it is known as ‘mal de Caderas’ or ‘Murrina’. These parasites have adapted to an entirely mechanical, non-cyclical mode of transmission by blood-sucking flies other than tsetse. Trypanosoma theileri is a stercorarian parasite of cattle which deserves greater mention. It was first reported by Theiler281 in the then Transvaal in 1903, and has since been found to occur in cattle throughout the world. It is transmitted by tabanid flies and is widely regarded as being non-pathogenic, but in certain circumstances it has been associated with disease.111, 280, 315
The following account of pathogenic trypanosomes deals only with tsetse-transmitted salivarian parasites, unless otherwise indicated. They are important as they cause severe disease. Human sleeping sickness is caused by Trypanosoma brucei gambiense and T. b. rhodesiense. Whilst these two subspecies do infect some domestic and wild animals, there are other, more significant pathogens of livestock which are found in three subgenera: Duttonella, Nannomonas and Trypanozoon (Table 12.1). The fourth salivarian subgenus, Pycnomonas, is represented by only one species, Trypanosoma suis, which is of little economic importance.
As a consequence of their pathogenicity and their complex and fascinating biology, the salivarian trypanosomes have been, and will continue to be, the subject of intense research. The remarkable alternate adaptations of these extracellular parasites to mammalian and insect hosts are reflected in morphological changes that are readily detectable by light microscopy. Bloodstream forms are trypomastigotes (Figure 12.2); from the posterior portion of an elongated body, some 8 to 35 μm long, arises a flagellum which extends anteriorly, and which is connected to the body by an undulating membrane. Beyond the anterior extremity of some species, the flagellum may extend free of attachment to the undulating membrane. The beating of the flagellum pulls the trypanosome forwards, imparting characteristic motility. Within the cell, in a posterior position and at the base of the flagellum, a kinetoplast is found, and a single nucleus is located almost halfway along the body. In the tsetse fly, trypomastigotes transform to epimastigotes (synonymous with the crithidial stage in the old terminology) in which the kinetoplast has migrated anteriorly, to a position adjacent to the nucleus (Figure 12.2). Differences in the morphology of the trypomastigote stages of the various species form the basis for differential diagnosis. The major characteristics are clearly seen in thin blood smears, stained with Giemsa’s, Leishman’s or other Romanovsky stains. Trypanosoma congolense (Figure 12.3a) and T. vivax (Figure 12.3b) are monomorphic parasites, whereas T. brucei is polymorphic (Figure 12.3c). Trypanosoma congolense is the smallest species and is between 8 and 20 μm in length.
It has no free flagellum, and the kinetoplast is usually subterminal and marginal.224 Trypanosoma vivax is between 20 and 26 μm long, and has a long free flagellum and a large, often terminal kinetoplast. The long slender form of T. brucei is characterized by a long free flagellum, the presence of a conspicuous undulating membrane and a sub-terminal kinetoplast. The posterior end is narrow and often pointed, and this form is between 23 and 30 μm long in contrast to the short stumpy forms which range on average between 17 and 22 μm. The short stumpy form has a well-developed undulating membrane and usually has no free flagellum. The lengths of the intermediate forms range from 20 to 25 μm. The kinetoplast of T. brucei is smaller than that of either T. vivax or T. congolense. Trypanosoma simiae is between 12 and 24 μm long and displays some polymorphism: most parasites are long stout forms, sometimes with a short free flagellum. Short forms of T. simiae resembling T. congolense and long slender forms also occur.111
The reasons for the cyclical changes from trypomastigote to epimastigote and back to trypomastigote, in the course of the trypanosome’s life cycle, have become clearer as a result of ultrastructural and biochemical studies. The morphological changes which are readily seen reflect the trypanosome’s adaptation to the different physiological environments encountered in mammalian and insect hosts. Ultrastructural, biochemical and immunological studies196, 299, 300 have not only revealed the reasons behind the cyclical changes in morphology, but have also elucidated the parasite’s survival tactics. Trypanosomes show remarkable adaptation. They survive not only in the turbulent bloodstream, where they face vigorous immunological assault, but they also withstand the digestive enzymes of the tsetse fly’s alimentary tract.
Trypanosomes are therefore robust organisms. The cytoskeleton of closely packed, parallel microtubules, strongly interconnected by associated proteins, is very resilient and helps to retain the body’s shape. The cytoskeleton supports the surface membrane (Figure 12.4 and 12.5), which, in mammal-infective stages, is coated with a surface glycoprotein some 12 to 15 nm thick. Within the cytoplasm of the cell are organelles, which are supported by intermediate filaments. 125 A nucleus, Golgi apparatus, rough and smooth endoplasmic reticulum, glycosomes, lysosomes, vacuoles and single mitochondrion are all present. Microtubules also delimit the entrance to the flagellar pocket, which is an invagination of the cell membrane from which the flagellum emerges (Figure 12.5). This pocket is a highly specialized area of the cell membrane at which endocytosis occurs, in a manner similar to that found in mammalian cells.305 Serum albumin, lipoproteins, iron-bound transferrin and other nutrients are endocytosed from the flagellar pocket by bloodstream trypanosomes.
The exchange of the contents of the pocket is assisted by the constant, pump-like beating of the flagellum, which is made up of an axoneme and paraflagellar rod, and which also provides the means of propulsion. It is ensheathed in surface membrane and is attached to the body by a series of junctions and numerous filaments.262 The beating of the flagellum draws the cytoplasm of the cell up into a series of transient crests which form the undulating membrane.300 At the base of the flagellum is the basal body which is associated with the kinetoplast. This latter structure consists of bundles of DNA fibrils located in a dilatation of the cell’s single mitochondrion.111, 300
In the mammalian host, the abundance of glucose as an energy source is associated with repression of the mitochondrion which lacks cristae and is tubular. Energy is derived from the metabolism of glucose to pyruvate.299
Table 12.1 Pathogenicity1 of salivarian trypanosomes to livestock
| TRYPANOSOME SUBGENUS | TRYPANOSOME SPECIES | CATTLE | GOATS | SHEEP | PIGS | HORSES | DONKEYS |
|---|---|---|---|---|---|---|---|
| Trypanozoon | T. brucei 2 | + | ++ | ++ | + | +++ | ++ |
| T. evansi3 | ++ | + | + | ++ | +++ | ++ | |
| T. equiperdum4 | - | - | - | - | +++ | ++ | |
| Nannomonas | T. congolense | +++ | ++ | ++ | + | ++ | ++ |
| T. simiae | - | + | + | +++ | - | - | |
| Duttonella | T. vivax | +++ | ++ | ++ | - | ++ | + |
| Pycnomonas | T. suis5 | - | - | - | ++ | - | - |
Notes
|
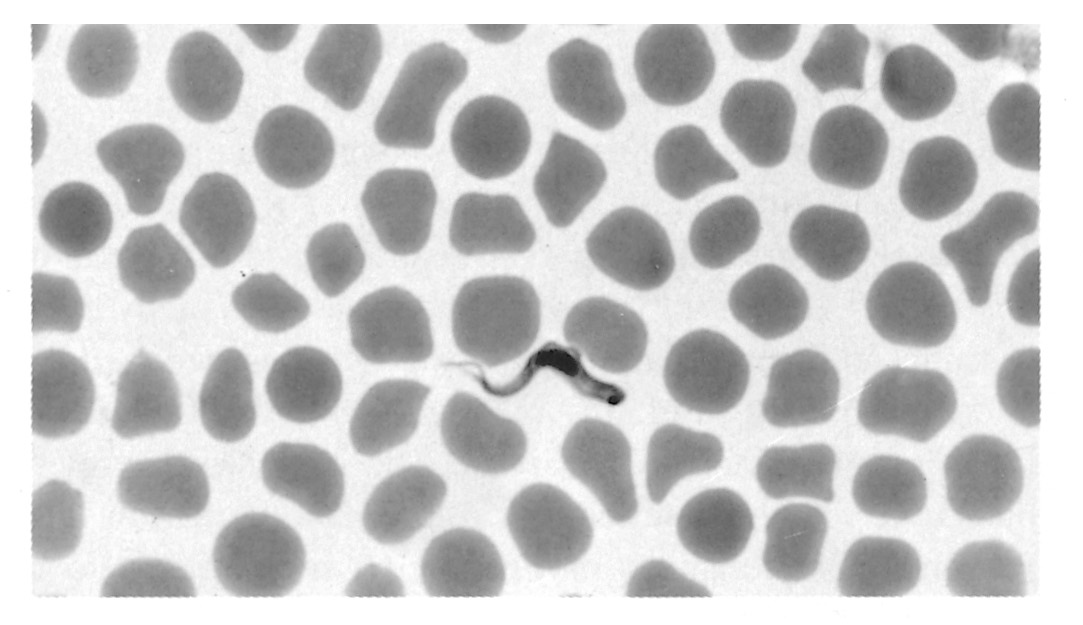
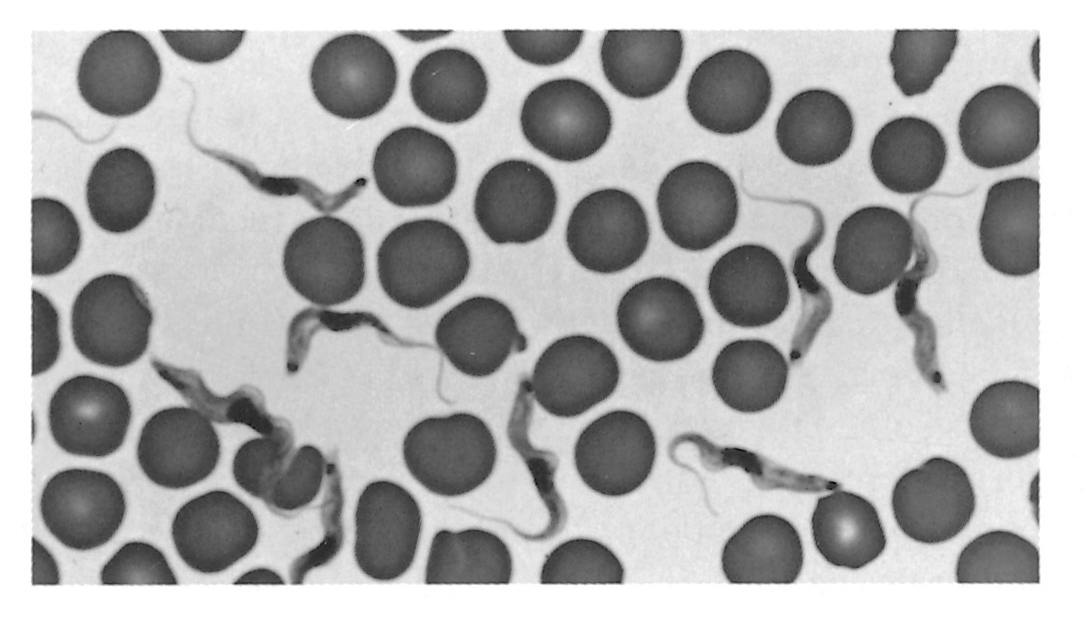
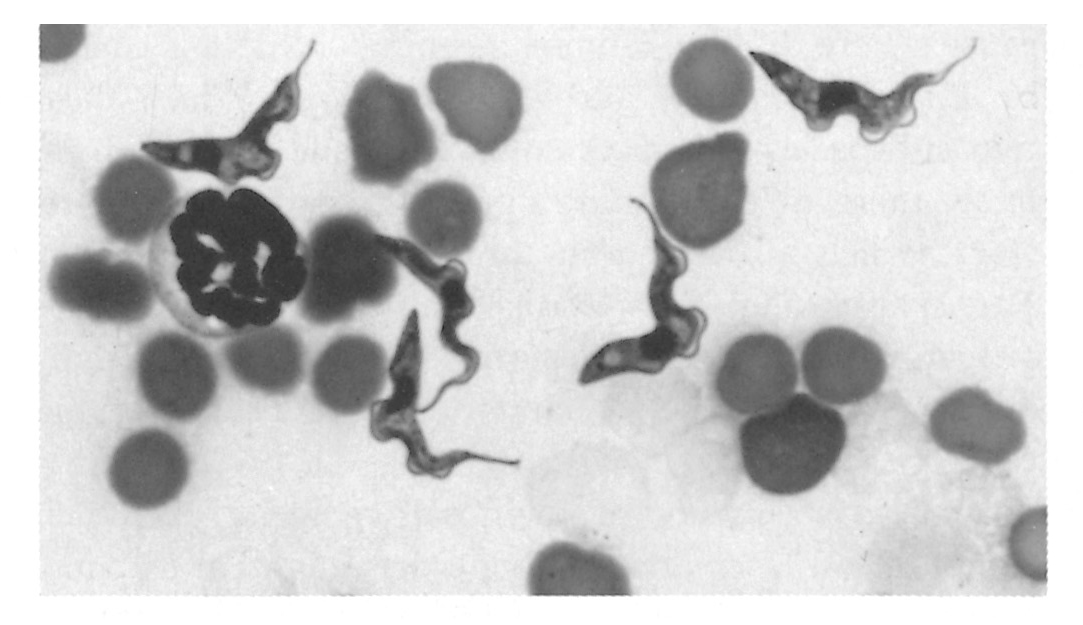
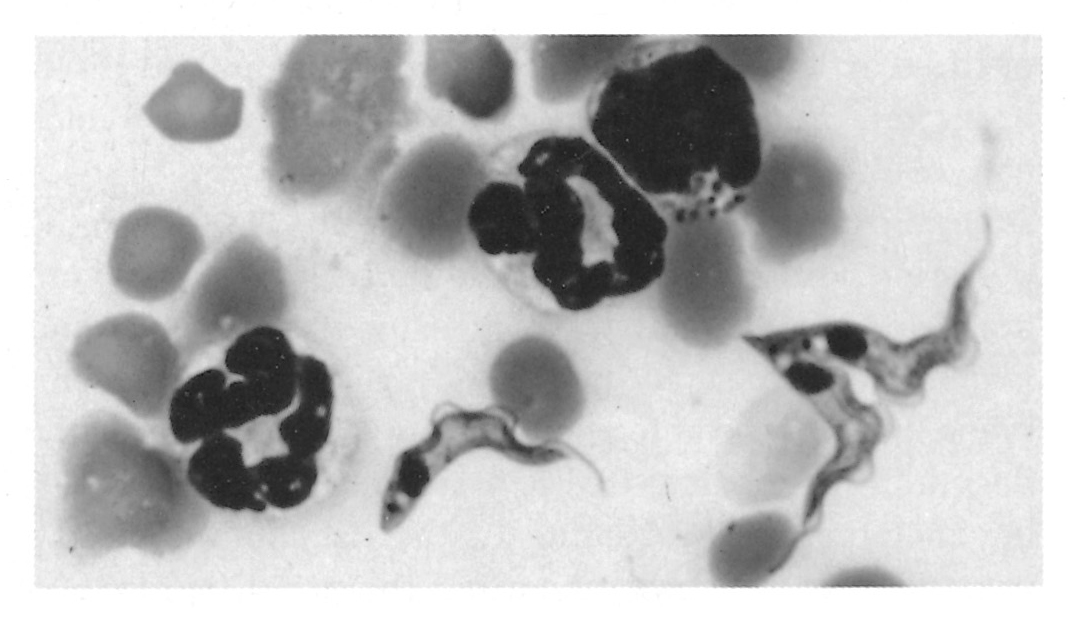
Figure 12.3 Trypanosomes in thin blood smears, x1 000 stained with Diff-Quick. a = Trypanosoma congolense: note absence of free flagellum; b = Trypanosoma vivax: note long free flagellum and large kinetoplast; c = Trypanosoma brucei: note polymorphism, prominent undulating membrane and free flagellum; d = Trypanosoma brucei dividing by longitudinal binary fission. (Unpublished photomicrographs by courtesy of Dr L. Logan-Henfrey, Laboratory for Research on Animal Diseases, PO Box 30709, Nairobi, Kenya)
However, the mitochondrion transforms in the tsetse fly, swelling and acquiring tubular cristae. Enzymes appear which oxidize proline, the main source of energy for both fly and trypanosome.299 This transformation is evident as the kinetoplast migrates and the trypanosomes assume the epimastigote form in the fly.
Trypanosomes reproduce by longitudinal binary fission, both in the bloodstream (Figure 12.3d) and in the fly, although a sexual process can apparently occur in the tsetse fly.132, 278 Multiplication in each host culminates in the presence of mature trypanosomes, which stop dividing and are pre-adapted to the conditions that they will encounter in the next cyclical host. As a tsetse fly takes its blood meal from an infected host it ingests trypanosomes. Pre-adapted parasites survive in the fly, but trypanosomes that are not metabolically adapted to the new physiological conditions die. The cycle of development of different species of salivarian trypanosomes is very similar, but there are important differences in their localization in the tsetse fly during their development (Figure 12.2). Trypanosoma vivax has the simplest migratory pattern; development occurs only in the proboscis and the pharynx (cibarium).111, 130 Bloodstream trypanosomes are taken up through the food canal of the proboscis, and the pre-adapted, late bloodstream forms attach to the labrum wall by means of the flagellum.88
Parasites that are swept to the midgut do not survive. In the proboscis the attached parasites lose their surface coats, transform to epimastigotes and multiply in the labrum. They then detach and move to the hypopharynx where, after re-attachment, they transform to metacyclics. The complete cycle takes 5 to 13 days, before glycoprotein-coated metatrypanosomes can be inoculated as the fly feeds.88 The development of members of the subgenus Nannomonas and Trypanozoon is more complex and goes through an immature or midgut stage. In the midgut of the fly, pre-adapted trypanosomes, or short stumpy bloodstream forms in the case of T. brucei, transform to procyclics, elongate and shed the glycoprotein coat. The coat is progressively replaced in a short time by a coat of procyclin, which probably provides protection against proteolytic enzymes of the midgut.250 The procyclic trypanosomes begin to metabolize proline as a source of energy for which they compete with their host, the tsetse. The transformation of bloodstream trypanosomes into procyclic or midgut forms within the fly’s midgut is a crucial first step in the establishment of a trypanosomal infection. Transformation proceeds rapidly in the posterior part of the midgut, the first procyclic forms appearing 11 hours after ingestion. 288 Factors known to influence this process include trypanolysins and trypsin or trypsin-like molecules in the fly’s midgut,126, 127 the type of host blood at the time of the infective feed,190, 194, 203, 246 and blood composition.93, 185, 230
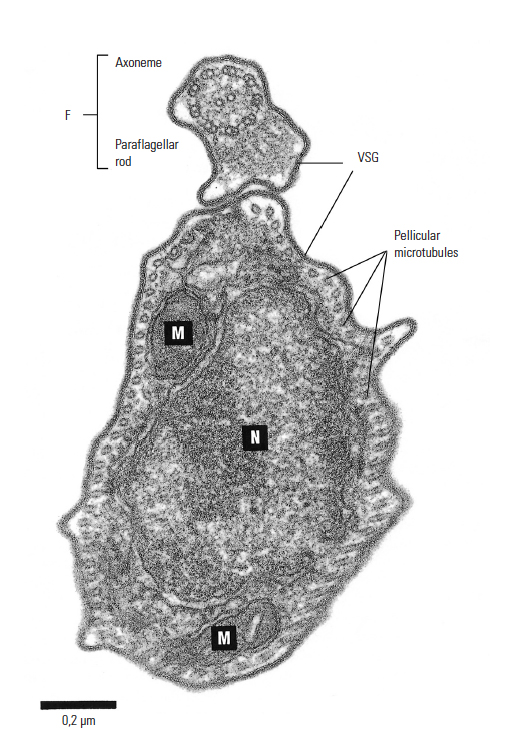
Figure 12.4 Electron microphotograph of Trypanosoma congolense: cross-section showing flagellum (F), nucleus (N), mitochondrion (M) and variable surface glycoprotein coat (VSG), x86 000. Bar represents 0,2 μm. (Unpublished electron micrograph by courtesy of Dr P. Webster, Yale University School of Medicine, Department of Cell Biology, New Haven, CT)
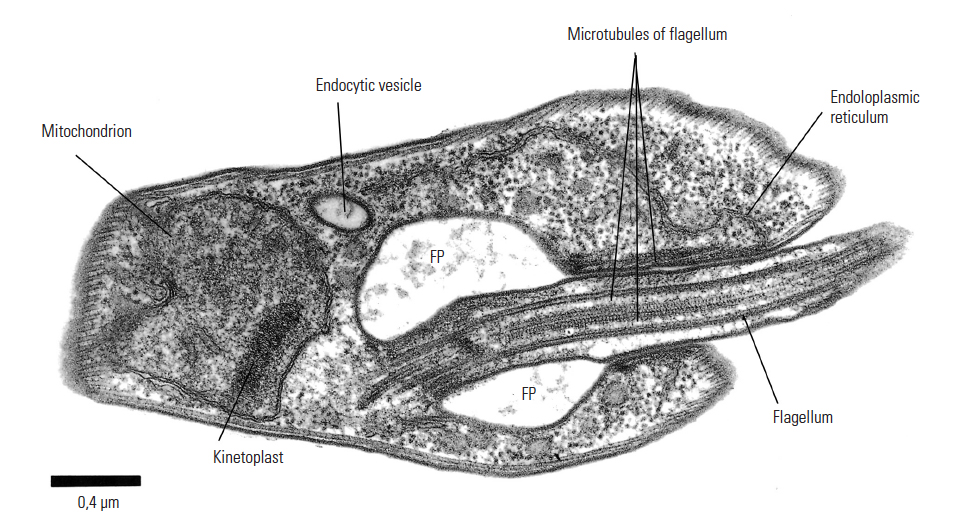
Figure 12.5 Electron micrograph of Trypanosoma brucei: section through the flagellar pocket (FP) region of the cell. Microtubules are longitudinally sectioned, x44 000. Bar represents 0,4 μm. (Unpublished electron micrograph by courtesy of Dr P. Webster, Yale University School of Medicine, Department of Cell Biology, New Haven, CT)
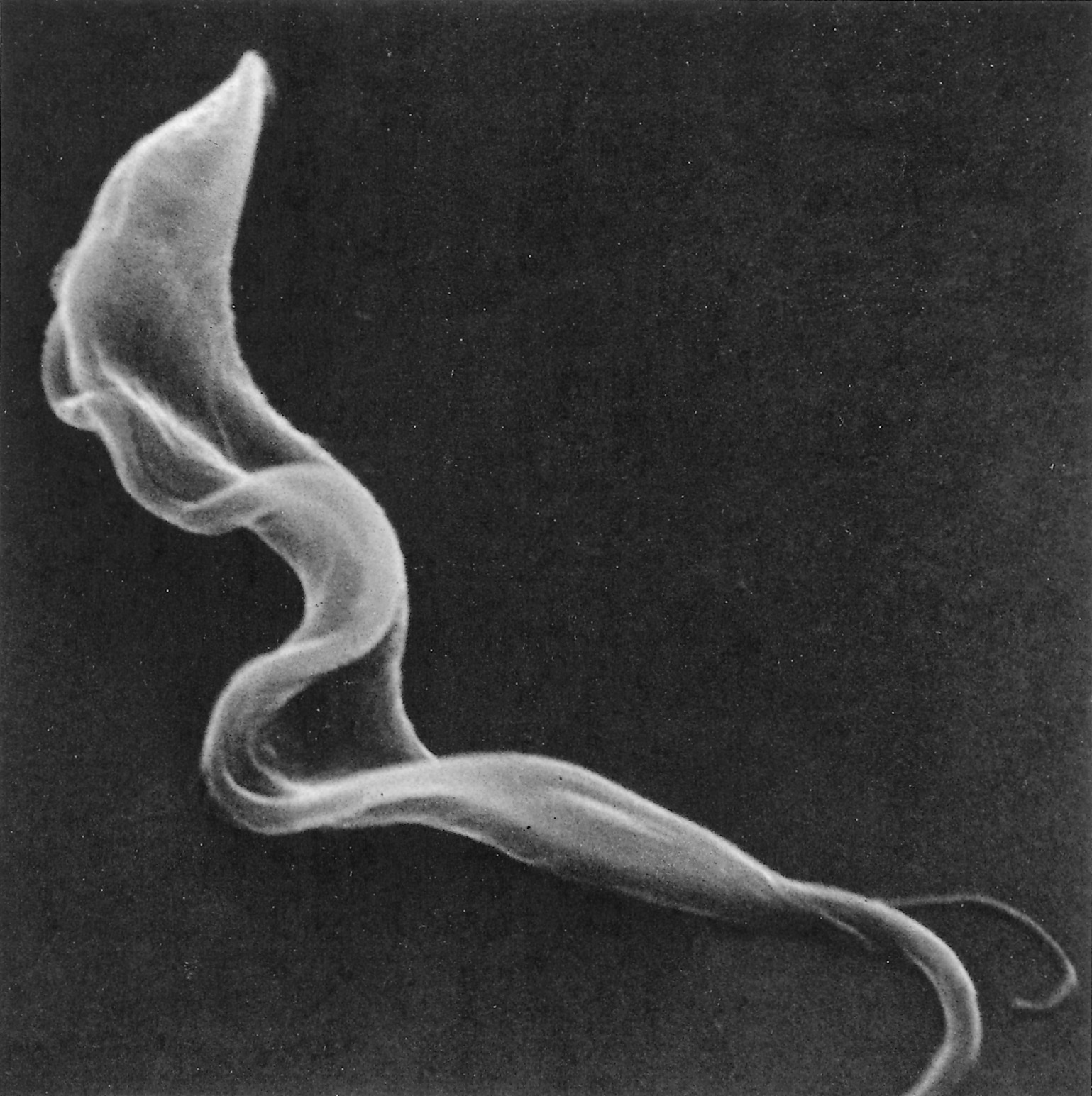
Figure 12.6 Scanning electron micrograph of an intermediate (bloodstream) form of Trypanosoma brucei from the blood of a mouse. Note the prominent undulating membrane, pointed posterior end and long, free flagellum. A ‘streamer’ or filopodium can also be seen. (By courtesy of Dr P. Gardiner and reprinted by kind permission of Vinand Nantulya and Parasitology Today)
The tsetse fly’s immune system also plays an important role. A humoral mechanism involving lectins is implicated in the establishment of trypanosome infections in Glossina. 186, 187 In invertebrates, lectins bind to specific carbohydrate groups on cell surfaces and may agglutinate certain cells. This may result in lysis and death of procyclic trypanosomes. 49, 119, 128, 129, 220 In the midgut, the titre of lectins and the degree of inhibition by lectins largely determine the ease with which trypanosomes in an infected blood meal establish a midgut infection.
The mechanism of maturation of a midgut infection is complex and, once established, it does not always progress to maturation. Before the infection is mature, procyclic forms transform into epimastigote and then to metacyclic forms. From the midgut, trypanosomes migrate to the mouthparts or salivary glands. The midgut procyclics are free swimming; they move to the ectoperitrophic space to form an actively dividing population.
They lose their glycoprotein coat and move forward to the proventriculus where they stop dividing. The proventricular ‘mesocyclic’ trypanosomes are longer than their procyclic precursors; they reinvade the endoperitrophic space and, in the case of membersof the subgenus Nannomonas, move via the oesophagus to the hypopharynx where they attach and complete their development to become coated metacyclics after 7 to 40 days. Members of the subgenus Trypanozoon migrate via the oesophagus, mouthparts and salivary ducts to the salivary glands where the parasites form flagellar attachments to the epithelial cells.
In the salivary glands the trypanosomes go through four stages of development. The mature, coated metacyclic trypanosomes undergo morphological and metabolic changes that pre-adapt the parasites for their life in a mammalian host. The development cycle of T. brucei takes from 17 to 45 days.175
As the infective tsetse fly feeds, metacyclic trypanosomes and saliva pass through the hypopharynx and are inoculated intradermally; it is here that infection is established. 3, 68, 79 Trypanosomes multiply in the skin, and may produce a chancre,248 which is a local skin reaction that develops into a raised, indurated, hot, painful swelling. The chancre may attain a diameter of 100 mm in 10 to 12 days, but regresses 10 to 15 days later.3 From the skin, the trypanosomes reach the blood via the draining lymphatics within a few days.3, 16, 71, 78, 168, 174 Trypanosomes multiply in the bloodstream, and although initially their low numbers make detection difficult, the generation time of only a few hours soon leads to high levels of parasitaemia. Trypanosomes may leave the bloodstream to reach various extravascular sites.
Trypanosoma congolense, for long regarded as a strictly intravascular parasite,116, 205 recirculates in lymph4, 71 and has been recovered from the central nervous system.184 However, this species is more commonly located in capillary beds where parasites attach to the endothelium.36, 166 Trypanosoma vivax shows a greater tendency to invade tissue and has been demonstrated in cerebrospinal fluid, perivascular spaces of the central nervous system, the aqueous humour of the eye, myocardium and in other tissues.183, 318
In contrast to T. congolense and T. vivax, infections with Trypanozoon species are characterized by generally low parasitaemias and a marked invasion of tissues.96, 117
Because of the greater accessibility of bloodstream parasites to investigators, this stage of the life cycle has been intensively studied. The ability of trypanosomes to establish prolonged infections is attributable to the phenomenon of antigenic variation.22, 64, 98 Each bloodstream trypanosome is completely clad in a dense surface glycoprotein coat298 (Figure 12.4 and 12.5), which consists of several million tightly packed molecules, each of which is anchored in the parasite’s surface membrane.250 Within a population of trypanosomes originating from a single infection, almost all bear the same glycoprotein coat and are thus of the same antigen type. As parasitaemia rises, a swift antibody response is elicited against the antigen type exposed on the surface of the bloodstream trypanosomes.
These specific antibodies attach to the surface glycoprotein and produce complement-mediated lysis of all trypanosomes of that antigen type. However, before antibodies reach trypanolytic levels, some trypanosomes — as few as one in 100 000—switch off the gene that controls the production of the initial surface glycoprotein and activate a gene that codes for a different protein.222 Trypanosomes which bear the new surface glycoprotein are of a different antigen type and are not destroyed by antibody against the first antigen type; they survive to produce another parasitaemic wave, which in turn is removed by antibody specific for that antigen type.
By this time a third variant has arisen, and, escaping the effect of host antibody, it survives to produce the next parasitaemic peak. This antigenic variation is the result of sequential expression of variable surface glycoproteins (VSGs) which constitute a repertoire of variable antigen types (VATs). Infections arising from a single trypanosome may have a repertoire of more than 100 VATs.222 Thus shielded from total destruction, trypanosome infections usually run prolonged courses, since each VAT is present for several days before being removed. Although there is a mixture of a small number of VATs within a parasitaemic peak, the sequence of expression of VATs tends to be quite stable in clonally derived trypanosomes.206 This imparts immunologically distinct characteristics to a strain of trypanosomes, the distinct strain being called a ‘serodeme’. In the course of successive parasitaemic waves, some trypanosomes stop dividing and transform to the pre-adapted form able to survive in the tsetse. This is most readily seen in T. brucei infections. The long, slender trypanosomes rapidly divide to produce parasitaemic waves, but some differentiate through intermediate forms to become non-dividing, short stumpy parasites. These pre-adapted parasites do not shed their surface coats.299 Precisely what induces transformation is not known, but it is thought to be related to the host’s susceptibility to infection and disease.24, 198, 263 Similar pre-adaptive changes have been found in T. vivax88 and are believed to be necessary for the cyclical development of T. congolense.
The survival of the non-dividing, pre-adapted trypanosomes in the bloodstream may be associated with the ability to shed the VSG coat. Also, the ‘flow’ of VSG molecules over the surface membrane enables VSG-bound antibody to be endocytosed in the flagellar pocket.300, 306 In addition to removing potentially harmful antibody and delaying complement- mediated lysis of the trypanosome, this process may also enable VSG to be recycled. The shedding of VSG by stumpy forms of T. brucei198 and the formation of ‘streamers’ or filopodia (Figure 12.6), consisting of coated membrane, 300 may represent a protective mechanism.290 The stiffer cell membranes of stumpy T. brucei parasites may also provide some protection against antibody-mediated lysis.299
After ingestion by the tsetse, pre-adapted trypanosomes shed the glycoprotein coat, transform, multiply and finally mature. Infective tsetse then transmit metacyclic trypanosomes to another host. Irrespective of the VAT of the bloodstream trypanosomes ingested by a fly, the metacyclic VATs of a serodeme are relatively constant.
After repeated tsetse-transmission of a single serodeme, the composition of metacyclic VATs is usually very similar. Although the metacyclic VATs of a serodeme are always similar in contrast to the large VAT repertoire of bloodstream trypanosomes, there is nevertheless antigen heterogeneity, even among the metacyclics within a single tsetse fly.206, 222 Many serodemes occur within a single species, each serodeme having its own VAT repertoire. The antigenic diversity within a species leads to the possibility of animals in a tsetse-infested area being exposed to a large number of antigenically distinct trypanosomes, but although trypanosomes within a species may be antigenically dissimilar, they are morphologically indistinguishable.
The characterization of trypanosomes for a long time relied on comparisons of their morphology, motility, host specificity, tsetse transmissibility and their development within the fly, but more recent characterization methods include isoenzyme typing, analysis of kinetoplast DNA by polyacrilamide gel electrophoresis, pulsed field gradient electrophoresis of chromosomal digests and DNA hybridization. 88, 91 In view of the great diversity which exists within a species, attempts have been made to standardize the nomenclature of distinct populations of trypanosomes.323 It has been recommended that the use of the term ‘strain’ should be avoided, but it is still widely used. In the context of veterinary medicine, certain strains of trypanosomes are recognized for their low or high virulence, the ease with which they are transmitted by tsetse, or for other broad characteristics which may not be entirely stable.
The sequel to infection with salivarian trypanosomes is not always disease. The outcome is determined by many factors, frequently related to the susceptibility of the host and the pathogenicity of the trypanosome. In the case of wild animals, a natural cycle of trypanosome transmission occurs which is not associated with disease. Similarly, in some breeds of domestic animals, infection with salivarian trypanosomes is tolerated, and host and parasite reach an equilibrium. Disturbance of the equilibrium may precipitate disease a long time after establishment of the infection. Thus, although the tsetse-transmitted trypanosomes are aetiological agents of African trypanosomosis, infection is not always synonymous with disease.
The occurrence of T. theileri in healthy cattle throughout the world111 exemplifies a well-developed host–parasite relationship. This stercorarian species is the largest trypanosome of cattle, and bloodstream forms reach a length of 100 μm. This stage is characterized by a long, thin, pointed posterior end, a long, free flagellum and a prominent undulating membrane. The large kinetoplast lies far from the posterior extremity, often in a marginal position. The nucleus is found midway along the body. However, these parasites are infrequently seen since infections are characterized by low parasitaemia. Within the bovine, T. theileri multiplies by binary fission in the tissues, mainly in the epimastigote form. When multiplying parasites are found in the bloodstream, polymorphic ‘immature’ forms are present.
When T. theileri is ingested by tabanid flies, a cycle of development occurs in the hindgut and infective metatrypanosomes are voided with faecal material. Several species of horseflies have been incriminated in the transmission of T. theileri (see Vectors: Tabanidae), which is mainly accomplished as metatrypanosomes penetrate intact oral mucosa.27 This occurs as cattle use their tongues to fend off biting tabanids and simultaneously ingest their faecal material. The contamination of broken skin with metatrypanosomes of T. theileri is a less important route of entry. The prepatent period ranges from four to six days,27 but detectable parasitaemia is short-lived and is not accompanied by clinical signs although infrequent reports incriminating T. theileri as a pathogen have been made.280 In the latter, concomitant diseases usually existed and probably accounted for clinical signs, reduced host resistance and a consequent rise in T. theileri parasitaemia.
Similar events occur in many wild animals which harbour tsetse-transmitted trypanosomes. Infected animals show no clinical signs,202 but when they are subjected to the stress of capture, for example, their immunity is reduced, parasitaemia flares up and clinical disease may be precipitated. The equilibrium can be restored by reducing the stress factors and by providing trypanocidal treatment. Losses in captured wild animals due to trypanosomosis may be reduced by providing trypanocidal treatment at the time of capture.
Epidemiology
The epidemiology of African trypanosomosis is almost entirely dependent on tsetse flies.136 African trypanosomes are well-adapted parasites of many species of wild animals, and sylvatic cycles of trypanosome transmission occur throughout the 10 million square kilometres infested by this unique vector. The distribution of tsetse in southern Africa is described in (Vectors: Tsetse flies), but within the general ecological limits of distribution the problem of trypanosomosis is not static. The 1990 outbreak of bovine trypanosomosis in the KwaZulu-Natal Province of South Africa28 testifies to the dynamic nature of the problem.
The natural hosts of salivarian trypanosomes usually show no clinical signs of infection, host and parasites being in equilibrium.202 The large numbers of naturally infected wild animal hosts constitute a huge reservoir of trypanosomes. Once infected, tsetse remain so for life and thus they too form a reservoir of infection. Consequently, when domestic animals are introduced into areas in which sylvatic cycles of trypanosome transmission occur, trypanosomosis always emerges as a serious disease.175 Wild animals are the natural hosts of T. brucei rhodesiense, the aetiological agent of human sleeping sickness in central, eastern and southern Africa.
Thus, people living and working in tsetse areas are at risk of contracting the disease, but for animal trypanosomosis to occur it is not always necessary for livestock to enter tsetse-infested areas; tsetse also move. The seasonal dispersal of fly, during the single rainy season in southern Africa, frequently results in an increased seasonal disease risk to livestock kept some distance from the primary tsetse habitat.292
Changes in land use may also alter the extent of tsetse infestation (see Vectors: Tsetse flies). The abandonment of cultivation, for various reasons, permits the regrowth of vegetation, which may then provide suitable tsetse habitats. Conversely, the intensive settlement and cultivation seen in some areas (throughout Malawi,55, 292 for example) destroy tsetse habitats. Although removal of tsetse should also remove the problem of trypanosomosis, livestock may have to graze some distance away from cultivated areas, along the edges of forest reserves, and may therefore still be exposed to tsetse fly challenge.
The epidemiology of tsetse-transmitted trypanosomosis is complicated and is well reviewed.174, 175, 254, 255, 324 Whiteside321 identified at least 18 major variables in the epidemiology of African animal trypanosomosis, which relate to the interactions of tsetse, wild hosts, livestock and their management, the trypanosomes and climatic conditions. The trypanosomal infection rate in tsetse is of prime importance. The ease with which infections develop in tsetse depends upon the fly’s vectorial capacity and specific factors related to the blood of host animals. However, within a particular tsetse population, the prevalence of metacyclic infections can vary significantly. For example, a recent study conducted in Zambia showed that the monthly total prevalence of mature trypanosomal infections in G. pallidipes could vary from 3,2 to 16 percent330 and is determined by factors such as the age of the tsetse population and the prevalence of trypanosomal infections in the host.
Tsetse can only transmit metacyclic trypanosomes if the flies live longer than the duration of the developmental cycle of a particular trypanosome species. The developmental period of trypanosomes varies and is parasite species-specific although a high degree of variability within a species has been observed.53 Generally speaking, the duration of the development of trypanosomes in tsetse increases with increasing complexity of the developmental cycle. The relatively simple cycle that T. vivax undergoes may last only for five days, compared with the 17 to 45 days required for the completion of T. brucei’s complicated cycle. Hence, the proportion of T. vivax infections in tsetse increases under adverse ecological conditions, when the fly’s shortened lifespan prevents the completion of longer cycles. Such adverse conditions often occur at the edge of fly-belts and result in the predominance of T. vivax infections in cattle.85, 159 A high proportion of T. vivax infections in cattle is, for example, observed at the edge of the fly-belts in western Zambia48 and in the Lianshulu/Mamilli area of Namibia’s eastern Caprivi.295
The prevalence of metacyclic infections in field-caught tsetse increases with the age of flies.107, 158, 257, 260, 331, 333 In the case of T. vivax, which does not pass through the tsetse’s midgut, the rate of acquisition of new infections with increasing age is often constant.332 Up to 100 per cent infection rates with T. vivax can occur in tsetse that repeatedly feed on T. vivax-infected hosts.39 For those trypanosome species that go through a procyclic stage in the midgut of tsetse (T. congolense and T. brucei), the relationship between age and infection is more complex and is affected by the reduced susceptibility of tsetse to infection as they age.
The prevalence of trypanosomal infections in tsetse is also affected by host preference. Two aspects are important in this context: firstly, there is a diversity of host preference among Glossina and, secondly, there is a variation among different species of hosts in their susceptibility to infection with the different species of trypanosomes.111 For example, G. austeni in Uganda, which has a preference for feeding on suids, has a low T. vivax infection rate, since pigs are refractory to infection with this species of trypanosome. In contrast, G. palpalis, which shows a preference for bovid blood meals, has been found to have considerably higher T. vivax infection rates.195 Tsetse species are often grouped according to those that feed mainly on suids, bovids, suids and bovids, and lastly, those that feed on most available hosts, including humans.307 However, host preference is not a rigid behavioural characteristic of a particular tsetse species. 29, 40, 135, 193, 249 The relative abundance of a host species may contribute to observed host preference. For example, within a country a tsetse species may feed mainly on suids in one locality and on bovids in another. This is the case in the Eastern Province of Zambia: on the plateau G. m. morsitans takes 75 per cent of its feeds on cattle,296 in the adjacent Luangwa Valley the same species takes most of its feeds from suids.264
An animal entering a tsetse-infested area risks becoming infected with potentially pathogenic trypanosomes that may produce disease. The degree of risk depends largely on the ‘challenge’. Although there is no adequate definition of ‘challenge’, 255 it is generally regarded as reflecting the number of infective bites that an animal receives in a given time. However, true challenge, or risk, is determined by the interaction of the number of infective tsetse bites, host preference, host susceptibility and the virulence of the parasite. Of particular importance is the relationship between absolute tsetse density and biting rate. A good understanding of this relationship is essential when predicting the impact of tsetse control interventions. Usually the challenge increases with the tsetse population density but even at low densities tsetse can still cause a substantial disease problem. This is partly attributed to the often observed, increased frequency with which flies that have metacyclic infections in their mouthparts probe.133, 197, 247
There are various reasons why a particular animal species may be subjected to greater challenge than another. The behaviour of a potential host can influence the ease with which a tsetse can engorge and this may contribute to observed preference. In Zimbabwe, the results of one study showed that the frequency with which blood meals were taken from cattle, sheep and goats by G. morsitans and G. pallidipes was 500:5:1 respectively. Observations on the relative attractiveness of these host species showed that the ratio of ‘probes:successful feeds’ was 3,8:1 for cattle, 28,0:1 for sheep, and 63,2:1 for goats respectively.243 Thus goats are attractive to tsetse but in response to the flies’ attempts to feed, they are vigorously defensive; they shake their heads, wrinkle their skin, stamp their feet and even attempt to bite the flies! Nevertheless, goats and sheep do acquire trypanosomal infections under natural conditions.21, 102, 271 Impala (Aepyceros melampus) behave similarly when tsetse attempt to feed. It can be surmised that sick animals, which are less vigorous, are at greater risk of being fed upon by tsetse. This may explain the increased feeding success of G. pallidipes on T. congolense-infected oxen.18, 19
Management practices may also alter the challenge to which livestock are subjected, and in this sense management is central to the epidemiology of trypanosomosis. The different management of calves and adult cattle can significantly reduce the level of challenge to which young animal are subjected.52 Observations on the grazing ranges of livestock in West Africa showed that while cattle foraged widely in tsetse-infested habitats, sheep, goats and donkeys remained closer to the villages. As a result, small ruminants and equids were less exposed to attack by tsetse than cattle.267 In southern Africa, the grazing patterns of communal cattle vary from season to season.261 In the cropping season (November to March) cattle are penned and herded away from the crops. In the early dry season (April to July), after the crops have been harvested, cattle are allowed to roam freely; they feed unattended mainly on crop residues. In the late dry season (August to October), the cattle have to move further afield to find grazing and browse, often entering tsetse-infested areas. Furthermore, the movement of animals from tsetse-free pastures to tsetse-infested watering points can create temporary challenge. Sedentary animals may face a seasonal increase in tsetse challenge as flies disperse during the rainy season. Even within the vicinity of a village, the actual challenge can be quite variable and can lead to differences in herd infection rates within a small area.302 Consequently, a thorough knowledge of local managementpractices is important to understand the epidemiology of nagana in a particular area.
Management also determines the general well-being of livestock. If, for example, animals are heat-stressed and their feed intake is therefore reduced, their resistance to trypanosomosis is also reduced, increasing the incidence of the disease.54 A careful distinction has to be made between an occurrence of this kind and increased challenge.
A major influence on the epidemiology of trypanosomosis is the use of trypanocidal drugs. Whilst permitting the use of tsetseinfested land, chemotherapy may alter the prevalence of trypanosome species in the area. In Zimbabwe, it was reported that the use of diminazene increased the prevalence of T. vivax in cattle.32 Furthermore, the repeated use of trypanocides may result in the emergence of drugresistant strains of trypanosomes, and the epidemiological picture then changes.259
Wild animal hosts of tsetse, such as kudu (Tragelaphus strepsiceros), warthog (Phacochoerus aethiopicus), bushbuck (Tragelaphus scriptus), bushpig (Potamochoerus porcus), African buffalo (Syncerus caffer), African elephant (Loxodonta africana) and square-lipped and hooked-lipped rhinoceroses (Ceratotherium simum and Diceros bicornis respectively), are vital to sylvatic cycles of trypanosome transmission and they form a major reservoir of infection. The increasing popularity of game ranching or farming frequently entails the translocation of these animals from tsetse habitats to tsetse-free areas. As a result, clinical disease may occur in animals normally regarded as ‘immune’, in the absence of the tsetse vector. There is also the possibility that trypanosomes could be mechanically transmitted from these species to livestock.
Whilst the bite of an infective tsetse clearly represents a trypanosomosis risk, infection does not always become established. Furthermore, even when an infection is established, disease does not always ensue. A variety of hostassociated properties modifies the outcome and further complicates the epidemiology of trypanosomosis. Estimates of the efficiency of transmission to susceptible cattle vary widely but field investigations suggest that it is very low (2,5 per cent).76, 191, 257
Wild animal hosts of tsetse and certain West African taurine cattle are tolerant of tsetse-transmitted trypanosomes. Carefully controlled experiments have shown that the trypanotolerance of N’Dama cattle and African buffalo is an innate characteristic.2, 106, 196, 208 This trait also occurs in some breeds of sheep and goats.102, 123, 217 Trypanotolerance is a much studied phenomenon.124, 198, 207, 210, 211, 252 Although the underlying mechanisms are incompletely understood, it is generally accepted that innate and acquired resistance, as well as environmental and management factors, affect trypanotolerance. Lower trypanosomal parasitaemias and less severe anaemia occur in trypanotolerant livestock than in trypanosusceptible animals.66 The epidemiological significance of this trait lies in the lower morbidity and mortality due to trypanosomosis in tolerant breeds. Nevertheless, even within a generally tolerant breed some individuals are less tolerant than others253 and tolerance may be affected by the degree of challenge.67 Similarly, within susceptible breeds individuals can be found that are apparently less susceptible to the effects of trypanosomal infection than most other animals. 50, 217, 231
Although trypanosome-infected animals may effect self-cure, the usual sequel to infection in tolerant animals is the establishment of a balance between host and parasite. If the host is stressed, the equilibrium is disturbed and a clinical episode of variable severity is precipitated. Stress takes many forms. Animals in late pregnancy or that are lactating are more susceptible to trypanosomosis. 175, 208, 233 Overwork also constitutes a stress, which, in infected trypanotolerant stock, may precipitate disease.175 This has serious consequences for animal traction and for hard-working bulls used in restricted breeding seasons in tsetse-infested areas. In a similar manner, intercurrent disease is stressful; trypanosome-infected animals with helminthosis or other diseases are more severely affected than those with either disease alone.104, 118, 175, 271
Adequate nutrition enhances the ability of infected animals to withstand the adverse effects of a trypanosomal infection. 145 Hence, trypanotolerance is greatly reduced by a low plane of nutrition, such as that available during the dry season.175 Inadequate nutrition results in generally lower average herd haematocrits, at all levels of disease prevalence. In the dry season, animals may also have to trek long distances to watering points. The combined effect of poor nutrition and increased exercise is often associated with an increased incidence of trypanosomosis in the dry season. This is not necessarily due to an increase in trypanosome transmission at this time of the year but could be explained by higher parasitaemias in animals with reduced tolerance. However, dry season grazing areas or watering points are often located in tsetse-infested areas, in which case the actual incidence can rise. Similarly, such a rise of incidence has been observed when cattle graze beyond the boundaries of a tsetse control area during the dry season 291
Age also has a significant effect on resistance to trypanosomosis. 211 It is widely recognized that cattle born in an infested area do not immediately succumb to disease, even though they acquire trypanosomal infections when young, whereas cattle brought into the area readily succumb. Three issues arise from this observation: firstly, young animals may be protected by maternally derived antibodies; 175, 211, 312 secondly, the ability of animals to acquire protective immunity is implied; and, lastly, young animals may be less attractive to tsetse and, thus, face a reduced challenge.
Despite the antigenic complexity of trypanosomes, infected animals do mount an immune response which, especially when supported by chemotherapy, can confer specific protection against homologous serodemes.97, 207, 329 Thus, within a defined area, animals may acquire protective immunity against locally prevalent serodemes. However, the movement of these animals to another area may expose them to different strains, or serodemes, to which they may succumb.175, 211 This type of tolerance requires continuous challenge and explains the low mortality of cattle in high challenge, endemic areas (such as the Eastern Province of Zambia) where trypanocidal drugs are used to prevent animals from succumbing to nagana.
The introduction of tsetse control measures in such areas would result in a loss of immunity that would render the population highly susceptible to infection and disease. If tsetse control measures break down, severe outbreaks of trypanosomosis would ensue, probably with high mortality, as happened following the disruption of tsetse control during Zimbabwe’s liberation war.156
Of great significance to the epidemiology of the disease is the nature of the parasite. Not all host species are equally susceptible to infection with each species of trypanosome (Table 12.1). Whereas T. vivax is a serious pathogen of cattle, pigs are highly resistant and, consequently, T. vivax infections are not seen in pigs. Conversely, T. simiae causes peracute and rapidly fatal disease in pigs, whereas cattle are apparently refractory to infection with this parasite.111 Within a trypanosome species are strains of differing virulence. Usually, T. vivax infections in domestic ruminants are prolonged with long aparasitaemic intervals. Occasionally, however, an acute haemorrhagic syndrome is seen in cattle infected with some strains of T. vivax.33, 179, 310 Infection with a single species of trypanosome can, therefore, produce quite different signs in the same host species.
An interference phenomenon also occurs, which either delays or prevents the establishment of an antigenically distinct trypanosome in animals with an existing trypanosomal infection.69, 70, 200 The mechanism of this interference remains unknown, but if the phenomenon occurs under natural conditions it would be epidemiologically important, since it could limit the number of infections to which an animal is subjected. However, by testing bovine sera with the highly sensitive polymerase chain reaction (PCR) technique it has been found that mixed infections are far more prevalent than had previously been appreciated.245
The occurrence of trypanosomal infections in areas apparently free of tsetse promoted the theory that infections can be maintained in nature by the mechanical transmission of trypanosomes by other haematophagous flies. In Malawi, a firm opinion developed that tsetse-transmitted trypanosomosis only occurred in close proximity to wildlife sanctuaries, where the flies were readily caught. Elsewhere, mechanical transmission alone was believed to occur.8 It is only relatively recently, with improved trapping methods that use attractant odours, that it has been possible to reveal the presence of Glossina at low population densities, which were hitherto undetectable. The recent discovery of G. pallidipes by these methods in the areas of Malawi thought to be free of tsetse55 may explain that part of the epidemiology of the disease in which mechanical transmission was thought to play a role.
Trypanosoma vivax has become established outside Africa, and especially in South America and Mauritius. In South America it is believed to be mechanically transmitted. 315 The cyclical development of T. vivax is confined to the mouthparts of tsetse flies and there is limited migration.
The close similarity of this mode of transmission to non-cyclical, mechanical transmission of T. vivax on the contaminated mouthparts of other haematophagous flies is widely assumed to have enabled this species of trypanosome to adapt readily to acyclical transmission. However, the possibility that cyclical development might occur in a vector, or vectors, other than Glossina spp., has not been completely eliminated. Nevertheless, there is some evidence that mechanical transmission of T. vivax can occur in areas adjacent to tsetse-infested areas. Reports of fulminating T. vivax infections which spread rapidly in the apparent absence of tsetse, leave little doubt that transmission can be mechanical, 46, 251 and the rapid spread of T. simiae within a piggery is believed to be due to mechanical transmission.
A wide variety of factors are involved in the epidemiology of trypanosomosis. It is their interaction which determines the clinical picture in an area, and, even though mathematical models have been developed to quantify the relative importance of these variables,256 much remains to be clarified and the models have yet to be applied to the field.
Pathogenesis
The precise pathogenesis of the trypanosomoses remains far from clear. Reviews of the subject164, 165, 274 highlight the complexity of this group of diseases: the various species of hosts involved differ in their susceptibility; there is great diversity among parasites within a species; and it is difficult to extrapolate the results obtained from experimental infections in rodents, ruminants and other animals, to explain the pathogenesis of natural infection.274 Three features — anaemia, tissue damage, and the suppression of immune responses205, 289 — dominate the pathology of trypanosomosis, and research aimed at elucidating the pathogenesis of the disease has largely addressed these areas.
Infection becomes established at the site of inoculation of metacyclic trypanosomes in the skin, where a chancre may form. Multiplication of the parasites induces an inflammatory response, the severity of which depends upon the size of the inoculum, the species of parasite and the breed and species of the host. In cattle, T. congolense is located in dilated lymphatics of the papillary dermis and hypodermis, in which the parasites form flagellar attachments to the endothelial cells.3 Dead and dying parasites are also present in the lesion, and various proteins and peptides have been isolated from the chancre. These may originate from either the host or the parasites.124 The chancre reaches a maximum diameter of some 100 mm, 10 to 14 days after an infective tsetse fly has fed, its development preceding invasion of the bloodstream by trypanosomes, and is accompanied by enlargement of the draining lymph nodes.3, 72, 79
Trypanosomes are detectable in the blood 13 to 16 days after an infective tsetse fly has fed.170 At this time the chancre begins to regress, and the characteristic series of intermittent parasitaemias begins. Parasitaemia rises and is accompanied by a febrile response, which is then followed by an aparasitaemic, afebrile period. The time between parasitaemic peaks varies from four to seven days in goats with T. vivax infections,183 to 12 days in cattle infected with T. congolense.82 In long-standing infections parasites may not be detected for months, and when parasitaemia does occur, it is often low grade and the animal is afebrile.
After an infection has become established, a protracted battle ensues as the parasite provokes an immune response, only to evade its full effect. Tightly clad in the thick, disposable protective glycoprotein coat, the trypanosome population is assured of continued survival in the face of the host’s vigorous defence. Anti-VSG antibodies destroy large numbers of trypanosomes, and dead and dying parasites and host cells are found extracellularly. These are removed by phagocytes and presented as antigenic evidence to cells of the lymphoid series. In the course of the contest, many harmful substances are released from both intact and damaged parasites and host cells. The destruction of large numbers of parasites releases lysosomal and other enzymes as well as structural proteins. Some of these enzymes have been identified and are thought to be directly harmful to the host. Many biological mediators, such as the vasoactive amines, are released from activated or damaged host cells. Activated macrophages release powerful cytokines,23 including interleukin-1 and cachectin (tumour necrosis factor), but their roles in the pathogenesis of trypanosomosis have yet to be investigated fully.
Much work has concentrated on comparisons of the responses of trypanotolerant and trypanosusceptible animals to infection. There is clear evidence that tolerant animals achieve better control of parasitaemia than do susceptible animals, and this ability is correlated with less severe anaemia and less severe disease.66 Anaemia is a cardinal sign of trypanosomosis in many domestic animals, and the aetiology is probably similar in all species. There is no single cause of the anaemia in trypanosomosis; the pathogenesis is complex and involves a variety of mechanisms, some of which are better understood than others.
Comprehensive reviews of anaemia in trypanosomosis209, 276 form the basis of the following account.
Trypanosomes release haemolysins and enzymes (proteases, phospholipases and neuraminidases) which directly damage red blood cell membranes. The fragility of erythrocytes is also increased by fever, and affected cells have a shortened lifespan. Erythrocytes are also damaged by trypanosomal antigens, which adhere to the red blood cell surface labelling it for removal. The attachment of antigenantibody complexes to red blood cell membranes also results in damage. Complement can be directly activated by trypanosomal VSG as well as being activated by antigenantibody complexes. The attachment of components of complement or complexed complement to erythrocytes produces damage and promotes erythrophagocytosis. Anaemia occurs largely because damaged erythrocytes are removed from the circulation by cells of the mononuclear phagocytic system (MPS) in the spleen, bone marrow (Figure 12.7), lungs and haemal lymph nodes.
To deal with the abundance of cellular debris and antigen-antibody complexes the MPS becomes greatly hyperplastic, which further enhances erythrophagocytosis. Intravascular haemolysis is thus not a prominent feature of trypanosomosis. A haemorrhagic syndrome is sometimes seen in acute T. vivax infections of cattle, in which sudden and severe thrombocytopenia occurs. This is associated with fulminating parasitaemia and extensive petechial and ecchymotic haemorrhages. The thrombocytopenia and resultant clotting defects, as well as the presence of circulating immune complexes, damage the capillary endothelium, leading to disseminated intravascular coagulation.
The removal of large numbers of red blood cells from the circulation occurs soon after the onset of parasitaemia, and produces a fall in the packed cell volume (PCV) — haematocrit. Packed cell volume is a reliable indicator of anaemia and is correlated with parasitaemia—the higher the parasitaemias, the lower the PCVs.
Accompanying anaemia is leukopenia, which possibly arises from direct inhibition of stem cell differentiation. Although there is an erythropoietic response to anaemia early in the course of infection, this seems to be impaired. Whereas anaemia is largely attributable to an increased rate of erythrophagocytosis early in infection, the anaemia of the late phase of infection has a different pathogenesis.
There are several possible sequelae to the early phase of infection, which depend largely on the nature of the parasite and the susceptibility of the animal. There may be spontaneous recovery or death, but very often there is a chronic phase which is characterized by infrequent, low-grade parasitaemias. Animals lose weight and condition and, as a result of dyshaemopoiesis, remain anaemic. Extensive haemosiderosis occurs as a result of erythrophagocytosis, and the trapping of iron in phagocytes is believed to contribute to the failure of erythropoiesis. Despite the apparent absence of parasites in the circulation, red blood cell destruction continues, and insufficient erythropoietic compensation results in persistent anaemia.
Trypanosomal parasitaemia correlates with complement activity in cattle. In the sera of N’Dama (trypanotolerant) cattle which develop lower parasitaemias than Boran cattle, complement activity is higher than in the sera of Borans infected with the same serodeme.125 Hypocomplementaemia is a frequent feature of the disease, with levels of component C3 being markedly reduced; this may be associated with reduced resistance to other infections.277 Various changes have been noted in the composition of leukocyte populations during the course of infection, but beyond the usual occurrence of leukopenia, other reported changes are equivocal.
The suppressive effect of trypanosomal infections on the immune responses of laboratory animals has often been demonstrated experimentally, but its significance in livestock has not been determined. Trypanosome-infected animals do mount protective, antibody-mediated immune responses against the parasites,198, 290 and both IgG and IgM are produced. The IgM levels are consistently elevated during infection and appear to be directed mainly against VSG antigens. Whilst some IgGs are directed against the common, somatic trypanosomal antigens, others appear to be directed against some of the host’s own cells. Anti-erythrocyte antibodies contribute to the anaemia of trypanosomosis and it is possible that a similar mechanism operates against leukocytes, contributing to leukopenia.209
Some animals are able to control parasitaemia quite effectively, and this appears to be antibody mediated, at least in part. Trypanotolerant Baoule cattle mount earlier and greater antibody responses to the first parasitaemic peak of T. congolense, compared with similarly infected Zebu cattle.2, 244 This ability to control the first parasitaemic peak is associated with the less severe anaemia occurring in trypanotolerant animals.
The antibody responses of trypanosome-infected cattle, sheep and goats to non-trypanosomal antigens are, nevertheless, depressed.173 The results of one investigation revealed that T. congolense-infected cattle produced poorer anamnestic responses to secondary vaccination with a polyvalent clostridial vaccine than did uninfected control cattle.112 On the other hand, cattle experimentally infected with T. congolense and vaccinated against foot-and-mouth disease were considered to have developed protective antibody titres.266 However, the effect of trypanosome-induced immunosuppression superimposed on the stresses of malnutrition, trekking, pregnancy or lactation may explain why affected animals frequently succumb to other disorders.
The precise aetiology of immunosuppression in trypanosomosis is obscure. It may involve B-cell mitogens, factors which block the release of antibody from plasma cells, reduced T-helper cell function, and depletion of lymphoid elements in the spleen and lymph nodes.290 Particles from dead and dying trypanosomes stimulate the MPS, and the cytokines released by cells of the MPS may contribute to the immunosuppression associated with the disease, as may products released by disintegrating trypanosomes.12, 290
There is a growing appreciation of the close functional relationship between the immunological apparatus and the endocrine system of the hypothalamo–pituitary–adrenal axis.17 Both systems are characterized by delicately balanced regulatory mechanisms, mediated by potent cytokines which have short half-lives. Interleukin-1 is a polypeptide released by macrophages and has a wide range of effects on many tissues. As well as stimulating T cells to produce interleukin-2, it is a potent pyrogen, stimulating the hypothalamus, the pituitary and the adrenal glands. Locally, interleukin-1 increases vascular permeability and is involved in the acute phase changes of the inflammatory process. The effects of trypanosomosis on the endocrine system have recently been demonstrated in cattle and goats.94 Trypanosome-infected animals show abnormalities of the thyroid gland, ovaries, testes, adrenal glands and pituitary. To what extent the changes are mediated by the direct action of the parasite, the host’s immune response or by imbalances in the endocrine system, remains to be determined precisely.
A parasite peptidase has been demonstrated in the plasma of T. congolense-infected heifers.151 The possibility exists that this enzyme might inactivate certain hormones or interfere with the host’s enzymes that are responsible for endocrine regulation. Such disturbance of the homeostatic mechanism would have serious consequences. In infected cattle, although levels of adreno-corticotrophic hormone are similar to those in uninfected controls, cortisol levels are lower, and this might indicate adrenal dysfunction or a decreased half-life of cortisol.125 The interpretation of research findings from laboratory-housed animals should always be made bearing in mind that chronic infections in stressed animals are commonly encountered in the field.
The pathogenesis of tissue lesions varies with the species of trypanosome. Trypanosoma congolense and T. vivax are mainly intravascular parasites; they induce changes in the endothelium of capillaries, and so indirectly cause damage to adjacent tissues. Trypanosoma brucei, on the other hand, has an affinity for tissues. Its presence in the extravascular compartment is associated with marked lesions in parasitized tissues.
The severity of endothelial injury also depends on the interaction of host and parasite. Trypanosoma congolense often attaches to erythrocytes and capillary endothelium. Damage to endothelial cells by parasite products, immune complexes, vasoactive amines and cytokines increases vascular permeability. In T. congolense infections a generalized dilatation of capillary beds, which alters the haemodynamics, is observed.164
The concomitant anaemia and more sluggish tissue perfusion affect the exchange of metabolites and are associated with intracellular oedema of capillary endothelial cells.221 Fibrinous microthrombi form in response to endothelial damage. These changes can be prominent in T. vivax infections, with which disseminated intravascular coagulation is more commonly associated.
Alterations to the microcirculation produce secondary degenerative changes in tissues. As capillary permeability increases, phagocytes and products of the parasite extravasate more readily and are responsible, in part, for some of the tissue lesions. Cytokines and parasite-derived substances, such as proteases, may directly injure host cells, an effect that is exacerbated by the formation of immune complexes in tissues. In the case of T. brucei, parasites localized in tissues cause mechanical disruption of host cells and probably also have some direct toxic effect on host cells, possibly mediated by their enzymes. The cellular response provoked by parasites within the tissues causes further damage, and auto-antibodies are also thought to play a role in inducing lesions.
The influence of all of these changes on the course of the disease depends upon their severity and upon the degree of impairment of the affected organs. An important feature of the pathogenesis of trypanosomosis is the effect on lymphoid tissue. As the disease progresses, the hyperplastic MPS grossly increases the volume of tissue in the spleen, lymph nodes and bone marrow. This hyperplasia of reticuloendothelial cells reduces lymphoid cell density, and eventual lymphoid depletion can occur.297
Chronic disease is associated with progressive emaciation and eventually cachexia. This is usually accompanied by low levels of parasitaemia, but the pathogenesis is poorly understood. The reduced lifespan of red blood cells, the increased catabolism of many proteins,277 and the associated negative nitrogen balance of clinically affected animals would appear to be largely responsible for the pitifully thin condition of animals suffering from chronic trypanosomosis.
Clinical signs, pathology and diagnosis in cattle
African trypanosomosis is transmitted by tsetse flies and is primarily caused in cattle by T. congolense and T. vivax. In endemic areas, the disease causes economic losses, but it can also occur in areas remote from tsetse infestations, as a result of the movement of previously infected animals which, when stressed, show clinical signs of trypanosomosis. Additionally, small numbers of infected tsetse flies, carried long distances by vehicles, can transmit trypanosomes in areas otherwise free of trypanosomosis. Recognition of the wide range of clinical signs associated with the disease is thus important, both within and outside endemic areas.
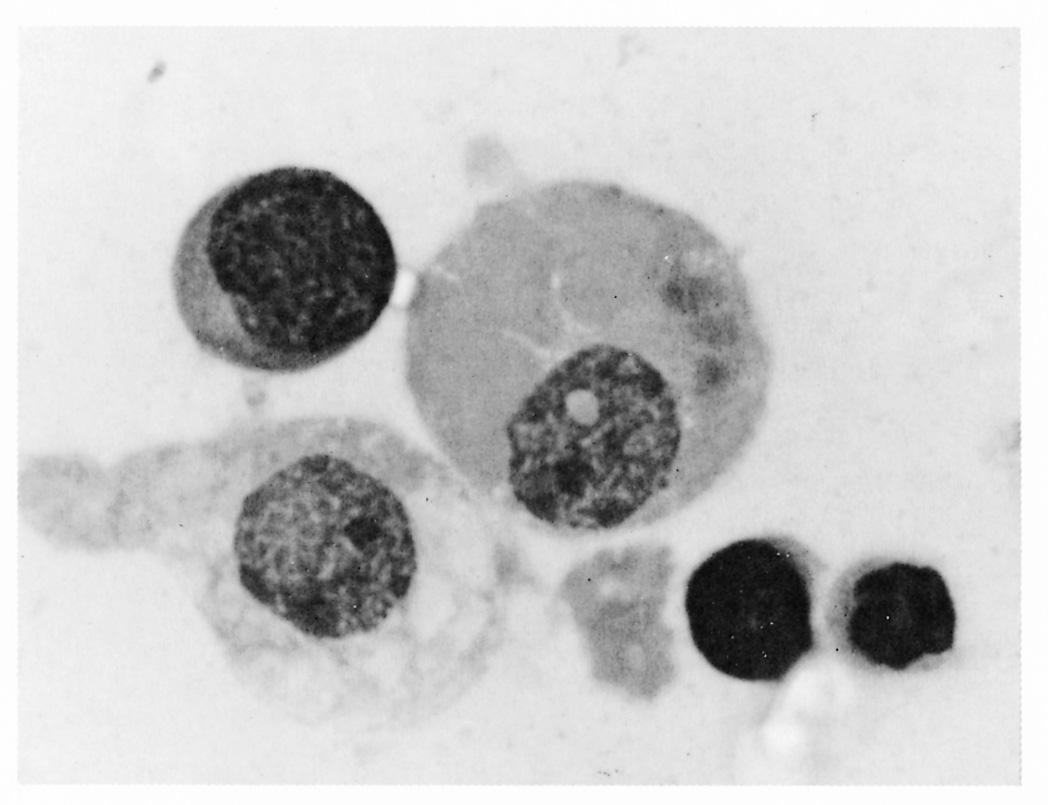
Figure 12.7 Bone marrow impression smear from a calf with an acute Trypanosoma vivax infection, showing a macrophage which has phagocytosed several erythrocytes x1 200. Stained with Wrights-Leishman. (Unpublished photomicrograph by courtesy of Dr L. Logan-Henfrey, International Laboratory for Research on Animal Diseases, PO Box 30709, Nairobi, Kenya)

Cattle living in tsetse-infested areas frequently have mixed trypanosomal infections which cause more severe disease than infections with a single species of trypanosome. Generally, in eastern and southern Africa, T. congolense is more pathogenic to cattle than T. vivax, and produces serious disease, whereas T. brucei is widely regarded as being of minor clinical significance.
Clinical signs
The course of disease due to infection with salivarian trypanosomes is variable and there are no clinical signs specific to bovine trypanosomosis. The manifestations of disease depend upon the degree of damage to specific organs and upon the degree of anaemia. Trypanosomosis in cattle may be acute, subacute or chronic; acute disease may be fatal after brief illness lasting two to six weeks, but chronic disease lasting many months or even years is more common.
Chancres are rarely seen in cattle with naturally acquired infections. The first signs of disease are due to the fever which accompanies the onset of parasitaemia. As parasitaemia falls, so does fever; the course of infection is therefore characterized by fluctuating parasitaemia and parallel fluctuations in body temperature. Intermittent parasitaemia and malaise are followed by increasingly severe clinical signs.
Acute trypanosomosis causes sudden reduction of milk yields and also abortion.116 Oxen, bulls and young stock with acute infections are reported to be suddenly ‘offcolour’. They are generally in good bodily condition but are dejected, their ears droop and they may walk stiffly. If animals are febrile at the time of examination, rectal temperatures may reach 39 °C in T. congolense infections, whilst in acute T. vivax infections they may be as high as 41 °C. Pulse and respiratory rates are raised in febrile animals and there may be piloerection. Conjunctival mucosae may be congested early in the acute phase, but later, as anaemia develops, pallor of the mucous membranes is evident.
Acutely affected animals quickly lose weight and body condition, although they continue to eat. They become weak, dejected and lethargic, often standing alone, away from the rest of the herd, not seeking shade. Excessive lachrymation is sometimes seen at this stage. After a short illness some acutely affected cattle become recumbent for a few days before death occurs. Corneal opacity of varying severity has been reported in some T. brucei and T. vivax infections.274 A proportion of acutely affected cattle gradually improve to enter the chronic phase of the disease. Occasionally, acute T. vivax infections induce a haemorrhagic syndrome,88 when animals bleed spontaneously from the ears, nose, mouth, anus and skin. Petechiae and ecchymoses may also be evident in the conjunctival mucosa.33, 179, 310
In contrast with the acute disease, subacute trypanosomosis is more common and runs a more prolonged course, with many animals apparently making a spontaneous recovery. However, they remain infected and frequently suffer from chronic trypanosomosis. In subacute cases (Figure 12.8), animals are intermittently ‘off-colour’, becoming weak and dejected, with drooping ears and flaccid tail. Within a few weeks they lose weight and condition, the coat becomes dull and a marked jugular pulse develops. On closer examination, although intermittent fever occurs, it is rarely marked; mucous membranes are pale, and pulse and respiratory rates are slightly raised. Superficial lymph nodes and haemal lymph nodes are frequently enlarged and are readily visible. Gradually, the frequency and intensity of parasitaemia decrease and fever subsides. Dependent oedema, particularly in the submandibular region, may develop. Thin, rough-coated, anaemic, lethargic cattle with generalized lymph node enlargement are said to have a ‘fly-struck’ appearance. Death may occur between four and six months after the onset of disease, but many animals make a gradual recovery, which is assisted by a good plane of nutrition.87
Chronic bovine trypanosomosis is by far the most common form of the disease in endemic areas. It may arise either as a result of partial recovery from acute or subacute disease, or from an initial subclinical or mild infection. Severely affected cattle may be extremely emaciated, having lost a large proportion of their muscle mass; they are often just ‘skin and bone’. Their general condition is very poor—the hair is sparse and the coat is rough, dull and staring. The skin is dryand scaly, and in long-standing infections the hair of the tail switch may be completely absent. The precrural lymph nodes and haemal lymph nodes are usually visible from a distance, and there is a marked jugular pulse. Although chronically affected animals continue to eat, they are weak and lethargic and are unable to keep up with the rest of the herd. Geophagia has been associated with bovine trypanosomosis, mainly in West Africa,108 but careful anamnesis often reveals that cattle with chronic trypanosomosis in other parts of Africa also have a tendency to eat sand or lick anthills. The chronic syndrome is characterized by infrequent, scanty parasitaemias, which are not accompanied by fever. Anaemia persists and is usually severe. Mucous membranes are very pale, the pulse is rapid, and breathing is laboured; in resisting restraint, the animal may collapse inrespiratory distress after brief exertion. The combined effect of anaemia, circulatory disturbance and myocardial damage frequently produces acute cardiac decompensation which leads to sudden death from congestive heart failure.
Although T. brucei normally produces mild or subclinical disease in cattle, signs of cerebral trypanosomosis arising from natural T. brucei infections have been described.313 One affected animal was described as a ‘loner’ — it would stray from the herd and become lost. This weak, emaciated animal displayed periodic generalized tremor, whereas another similarly affected animal in good condition showed hyperaesthesia and agitation.
Whilst for descriptive purposes it is convenient to classify trypanosomosis as being acute, subacute or chronic, the course that an infection actually follows is modified by many factors. A subclinically affected animal may, when stressed, suffer an acute clinical episode, while acutely affected animals may make a gradual recovery only to remain chronically affected. Yet other animals may effect an apparent self-cure, whilst continuing to harbour trypanosomes. The occurrence of concomitant disease frequently masks the underlying trypanosomal infection, but on occasions may exacerbate the clinical signs of trypanosomosis. The immunosuppression induced by trypanosomosis renders infected stock more susceptible to other diseases. In many cases, the underlying cause of disease or death may be concealed by the presence of the more readily detectable, secondary infection, such as anaplasmosis, helminthosis or lumpy skin disease. The consequence of this is that the true extent of trypanosomosis is often underestimated.
In endemic areas, trypanosomosis in cattle is a herd problem. The general condition of the herd is poor, although some individual animals are in surprisingly good condition. Trypanosomosis causes serious reduction in productivity, and careful analysis of records reveals its impact on fertility, which is reduced in both males and females.121, 131, 236 In addition to their being generally weak, the semen quality of affected bulls is reduced.265 Infected cows lose weight and their oestrus cycles become irregular andmaycease altogether.162 Thismay result in a significant reduction in calving rates. In the Eastern Province of Zambia, for example, the calving rate in a trypanosomosis endemic area is 44,1 per cent compared to 60,4 percent in an adjacent tsetse-free area.61 Trypanosomal infection during pregnancy may induce abortion,116, 131, 236 but when pregnancy is carried to term, the birth weights of calves are low.63, 234 Trypanosome-infected young stock grow poorly; they show signs of ill-thrift and remain stunted. The compounded losses from lower fertility and milk yields, increased mortality and reduced growth rates are reflected in low herd performance and often in decreasing herd size. A composite productivity index incorporating these parameters has been found useful when assessing the impact of bovine trypanosomosis and its control.182
The clinical signs of trypanosomosis tend to be more severe in exotic breeds of cattle than in the local breeds which, in part, reflects the adaptation of local breeds to their environment. Additionally, exotic breeds are often under greater production stress, which further exacerbates the disease.
Trypanosoma theileri is not considered to be a pathogen, 274 although high parasitaemias can occur in cattle with intercurrent disease.280 There are several reports associating T. theileri infection with such clinical signs as anaemia, diarrhoea or reduced milk yields, but conclusive evidence of the role of this species of trypanosome in the pathogenesis of these disorders is lacking.
Pathology
The PCV begins to fall with the onset of parasitaemia and fever. From an initial value of 0,32 l/l it may be reduced to 0,20 l/l or less in a matter of a few weeks. The early erythropoietic response to anaemia209 is usually normochromic and normocytic, but an early macrocytic response may occur. In chronic disease there is a microcytic anaemia, associated with hypoferraemia and dyshaemopoiesis due toiron trapping. Initial leukopenia also occurs and may be followed by leukocytosis. Thrombocytopenia is a constant feature of trypanosomosis and may account for the clotting defect that occurs in the T. vivax-haemorrhagic syndrome. 179, 209/ Hypocomplementaemia is also a constant finding in trypanosomosis,277 which is correlated with the level of parasitaemia;125 complement levels are higher in cattle that have lower trypanosomal parasitaemias. The levels of other serum proteins also alter in the course of trypanosomosis. Albumin is decreased and globulin levels are frequently raised.86, 277 Other changes also occur in blood chemistry — blood chloride levels rise and the calcium-tophosphorus ratio in infected adult cattle is significantly reduced. 82 More recently, attention has been directed to the effect of trypanosomosis on hormone levels. Results indicate that although plasma cortisol levels of infected cattle are reduced, parasitaemia appears not to alter levels of plasma adrenocorticotrophic hormone.125
The gross pathology of affected animals varies with the duration and severity of the disease, and many organs and tissues are affected. Acute infections may end with the death of an animal still in good bodily condition. In such cases the carcass is well-fleshed, although the mucous membranes may be pale. The overall appearance of the carcass is one of paleness, and petechial and ecchymotic haemorrhages are frequently present, especially on serosal surfaces. In acute T. vivax infections, there are extensive haemorrhages, resembling those seen in septicaemic conditions: large, epiand endocardial haemorrhages are present, in addition to those on serosal surfaces.
Lymph nodes are enlarged and, when incised, are oedematous and often have a dark pigmented medullary area. Typically, the spleen is greatly enlarged and dark red. Excessive peritoneal fluid is present which is often bloodtinged. Acute T. congolense infections can produce extensive oedema which is most noticeable in the perirenal and mesenteric fat, and the abomasal mucosa.164 Trypanosoma brucei may cause keratitis, although this is rare in cattle.
The gross pathology of chronic trypanosomosis is characterized by cachexia and anaemia. The coat is dry and dull, the skin is scaly and inelastic, the eyes are sunken, and the skeletal muscles are atrophied. There is hydrothorax, hydropericardium and ascites, and the carcass, in general, appears oedematous. Residual fat around the heart and kidneys is gelatinous. Lymph nodes, haemal nodes and spleen are generally enlarged and have an appearance similar to that seen in acute trypanosomosis, but after prolonged disease there may be splenic atrophy. Haemorrhages are not a feature of chronic trypanosomosis. The heart is enlarged and flabby, the liver swollen and pale. Whereas red marrow is present in the long bones in acute trypanosomosis, all the bone marrow is yellow and gelatinous in the chronic disease.
Gross changes are not always obvious, but microscopic lesions are extensive. These are found particularly in the cardiovascular system and lymphoid tissues, and there is widespread infiltration of many organs by mononuclear inflammatory cells. Dilation of the microvasculature and oedema and structural changes in vessel walls are usually seen, especially with T. congolense infections. Trypanosomes are commonly present in the lumens of blood vessels, and, in the case of T. vivax, parasites may be associated with fibrinous thrombi in the larger blood vessels. In some acute cases, there is disseminated intravascular coagulation, when microthrombi may be associated with haemorrhage and ischaemic necrosis. A constant feature of trypanosomosis is initial hyperplasia of the lymphoid tissues and of the mononuclear phagocytic system. In the cortical area of lymph nodes and white pulp of the spleen, germinal centres form early in the disease, although in chronic infections lymphoid tissue becomes depleted. 205 The expansion of the mononuclear phagocytic system leads to increased erythrophagocytosis and there is widespread haemosiderosis.
Monocytes and lymphocytes aggregate in the microvasculature together with trypanosomes. Invasion of tissues by the parasites leads to infiltration by mononuclear cells, which is mainly perivascular. In the late stage of acute trypanosomosis, severe anaemia and extensive vascular damage produce degenerative changes in the tissues, but these are less serious than are the changes that arise from cellular infiltration. Since T. brucei is more invasive, it provokes considerable cellular infiltration. The heart is commonly affected, and macrophages, lymphocytes and plasma cells infiltrate the myocardium. Degenerative changes and focal necrosis of myocytes and, in long-standing infections, fibrosis are evident in the heart. Infiltrations also occur frequently in other organs such as the pituitary, thyroids, adrenals, kidneys and gonads. Congestion and interstitial oedema also occur, and, when the substantia propria is affected, corneal opacity then develops. Severe degenerative changes occur in the thyroids82 and a reduction in erythroid progenitor cells of the bone marrow has been reported.209
Diagnosis
The specific diagnosis of trypanosomosis is notoriously difficult. Not only are there no specific clinical signs, but the intermittent and frequently low parasitaemias make detection of the parasites difficult.
Furthermore, infection is not synonymous with disease; many subclinically infected animals live in delicate balance with potentially pathogenic trypanosomes. An element of clinical judgement is therefore necessary when making a diagnosis of trypanosomosis. Several authors51, 148, 223 have compared the different methods for the parasitological diagnosis of animal trypanosomosis. Important considerations are that the number of detectable parasites is not necessarily related to the severity of the disease and, in endemic areas, the disease is a herd problem. The detection of infection in a few clinically affected cattle warrants careful examination of the entire herd.
Economic principles, the availability of expertise and the diagnostic requirements will guide the choice of a particular diagnostic test. The diagnostic method will differ between situations, depending, for example, upon whether species-specific diagnosis is required or whether surveys are conducted simply to determine the presence or absence of the disease. Often a combination of diagnostic tests is needed to obtain the required results.
Although in many tsetse-infested areas diagnostic facilities are not readily available, clinical signs of trypanosomosis are well recognized. Farmers and veterinary personnel commonly resort to treatment of sick animals, and use the response to therapy for retrospective diagnosis. In such areas, a history of the presence of tsetse flies and the use of trypanocidal drugs, when considered with presenting clinical signs, are sufficient to make a tentative diagnosis. However, the presence of concurrent disease may mask trypanosomosis and complicate the clinical picture. Thus, the only way to confirm diagnosis in clinically infected animals is to demonstrate and identify the parasites in body fluids.
The parasite detection methods are highly specific but their diagnostic sensitivity (the proportion of infections that the methods detect) is relatively low. This is especially the case when results are considered for an individual animal rather than in a herd. As a result, the apparent prevalence of trypanosomosis determined by parasitological diagnostic tests is an underestimate of the true parasitological prevalence. This is a problem in areas where the disease is present at low prevalence or is seasonal, or when attempting to confirm the absence of the disease in a particular area.
The body fluid most commonly examined is blood, either capillary blood from the tip of the tail or venous blood from an ear vein or from the jugular vein. Lymph, aspirated from a punctured superficial lymph node (usually the prescapular), provides useful supplementary diagnostic material. Whereas cerebrospinal fluid is routinely examined for the presence of trypanosomes in the diagnosis of human sleeping sickness, this has little practical use in veterinary medicine.314
Wet blood smears are prepared by placing a drop of blood (about 2 μl), taken directly from a punctured ear vein or the tip of the tail, onto a clean, dry, grease-free slide. This is immediately covered with a cover slip and examined microscopically as a fresh, wet preparation with a ×25 or ×40 objective lens. Approximately 50 to 100 fields are examined. Live, motile trypanosomesmaybe seen as they bore their way between blood cells (the name ‘trypanosome’ is derived from the Greek ‘trypanon’, a borer, and ‘soma’, meaning body). The wet blood smear method is simple, inexpensive and gives immediate results. It can be used to examine a large number of animals. Depending on the trypanosome’s size and movements a presumptive identification can be made of the trypanosome species involved. However, the diagnostic sensitivity of the method is low and depends to a large extent on the examiner’s experience and the level of parasitaemia. Sensitivity can be improved significantly by lysing the red blood cells before examination using a powerful haemolytic agent such as sodium dodecyl sulphate (SDS).228
More commonly, for routine diagnosis in veterinary practice, thick and thin smears of blood are prepared. A drop of blood taken directly from a punctured blood vessel in the ear or tail tip is placed on a glass slide; a thick and a thin blood smear can then be made on a single slide. Lymph smears are prepared in a similar manner. The smears are kept dry and protected from dust, heat, flies and other insects. Alternatively, under dusty field conditions, venous blood can be taken into tubes containing an anticoagulant, such as heparin or EDTA, and blood smears can be made subsequently. The dry smears are stained with Giemsa’s stain, thin smears after having been fixed, and examined under a ×50 or ×100 oil-immersion objective lens. Although it may be necessary to examine up to 700 fields to find a single trypanosome,51 the unfixed de-haemoglobinized thick smear allows approximately 120 times more blood to be scanned than a thin smear148 and, thus, has higher diagnostic sensitivity than the thin smears. Trypanosomes are easily recognized by their general morphology but may bedamaged during the staining process. This makes it difficult to make a species-specific identification on thick smears. The thin smear, on the other hand, permits accurate speciation of the parasites. However, due to the staining process, results are delayed.
The probability of detecting trypanosomal infections in a sample of infected animals can be improved by increasing the volume of blood to be examined and concentrating the trypanosomes. This is achieved by collecting fresh blood (about 70 μl) in a heparinized capillary tube and separating the different components of the blood by centrifugation, according to their specific gravity. Trypanosomes are concentrated in the plasma/white blood cell interface or buffy coat. The buffy coat is examined microscopically whilst slowly rotating the capillary tube (microhaematocrit centrifugation technique or Woo-method).330 Trypanosome movement can first be detected using the ×10 objective lens with reduced condenser aperture; the trypanosomes can be seen more clearly using a ×40 objective lens, preferably of a long working distance (allowing considerable depth of focus through the capillary tube).
The microhaematocrit centrifugation technique is more sensitive than the direct examination techniques but identification of trypanosome species is difficult. Alternatively, the buffy coat and the uppermost layer of red blood cells can be extruded onto a clean microscope slide and covered with a cover slip (buffy coat technique or Murray method212). Approximately 200 fields of the preparation are examined microscopically for the presence of motile trypanosomes with a dark-ground/phase-contrast illumination and a ×40 objective lens. Trypanosome species can be identified by reference to the following criteria:
- T. vivax: Large, extremely active, traverses the whole field very quickly, pausing occasionally.
- T. brucei: Various sizes, rapid movement in confined areas.
- T. congolense: Small, sluggish, adheres to red blood cells by anterior end.
- T. theileri: More than twice the size of pathogenic trypanosomes.
Since the specific gravity of T. congolense is similar to that of red blood cells, these parasites are often found below the buffy coat within the uppermost layer of red blood cells. The two concentration methods are the most sensitive for detecting T. congolense and T. vivax infections.238 The sensitivity of the concentration methods can be further improved by using the buffy coat double centrifugation technique.153 A total amount of 1 500 to 2 000 μl of blood is centrifuged after which the buffy coat is aspirated into a microhaematocrit capillary tube and centrifuged again. The buffy coat is then examined as previously described. A modification of the microhaematocrit centrifugation technique is the Quantitative Buffy Coat (QBC) method.14 This method is based on expanding the buffy coat by inserting a cylindrical float in a capillary tube that is precoated with acridine orange. The method has been used to diagnose T. b. gambiense infections but is too expensive for routine use in the diagnosis of animal trypanosomosis.
The microhaematocrit centrifugation and buffy coat techniques give immediate results and can be used for screening large numbers of animals. They require, however, specialized equipment and an electricity supply which makes the tests more expensive compared to the examination of the wet blood film, but this is compensated for by the increased diagnostic sensitivity; furthermore, haematocrit readings may be obtained (see below). Both parasite concentration techniques rely on the detection of live, motile trypanosomes. Since trypanosomes are heat sensitive, samples collected in capillary tubes should be kept cool; overheating of the microhaematocrit centrifuge should be avoided. Compared to the microhaematocrit centrifugation technique, the buffy coat technique has the added advantage that preparations can be fixed and stained for more accurate species identification and for retention as a permanent record.
The microhaematocrit centrifugation and buffy coat techniques are particularly useful in that the haematocrit or PCV can be assessed after centrifugation, by placing the microhaematocrit capillary tube in a haematocrit reader. The PCV of venous blood is a useful indicator of anaemia, which is one of the most typical signs of trypanosomosis in domestic animals.209 The PCV of individual animals and the average PCV of herds can be determined. In individual animals, haematocrit readings supplement parasitological diagnosis. In trypanosomosis-endemic areas, parasitologically negative animals that have low haematocrits are regarded as having trypanosomal infections. In surveys todetermine the presence or absence of trypanosomosis, samples collected from animals with low haematocrits can be investigated more thoroughly to increase the probability of detecting a trypanosomal infection. In parasitologically positive animals, the haematocrit indicates the severity of disease. The decision of the cattle owner to administer a trypanocidal drug is more often based on the effect of anaemia on the animal’s performance rather than the confirmed presence of infection. At the herd level, the haematocrit profile or the herd average PCV is a useful indicator of infection and herd health. However, other factors such as nutrition and fasciolosis may also cause anaemia on a herd basis. Therefore, it is important to establish the haematocrit profile of trypanosomosis-negative herds before relying on it as an indicator of trypanosomosis in a herd. Because of the low sensitivity of parasitological diagnostic tests, baseline PCV-profiles are best obtained from herds of animals that do not have anti-trypanosomal antibodies (Figure 12.9). In the absence of PCV-profiles of negative herds, comparisons of herd average PCVs at various distances from the source of infection — along a transect — can give useful information on the variation in disease challenge (Figure 12.10). In trypanosomosis endemic areas, herd average PCV is significantly correlated with parasitological prevalence (Figure 12.11). The slope of this relationship is determined by the effect of disease prevalence on herd average PCV and may be used as an indicator of the impact of trypanosomosis on herd health.
The subinoculation of blood into rodents, usually mice or rats, allows a greater proportion of T. brucei infections to be detected than by direct examination of the buffy coat. For practical reasons, subinoculation of blood into laboratory or other animals is not used as a routine diagnostic procedure. The method is expensive and diagnosis is not immediate. Furthermore, since rodents are refractory to T. vivax and not all T. congolense and T. brucei infections become established in the new host, even this method has serious limitations. Mixed trypanosomal infections may alsoremain undetected.
A procedure for the in vitro cultivation of T. brucei from the blood of infected animals has been described but success has been variable. Moreover, the method needs sophisticated equipment, yields results only after a considerable delay and is certainly not suitable for widespread use.
Figure 12.9 Herd average haematocrit profiles of anti-trypanosomal antibody negative herds and parasitological positive herds (Eastern Province, Zambia)
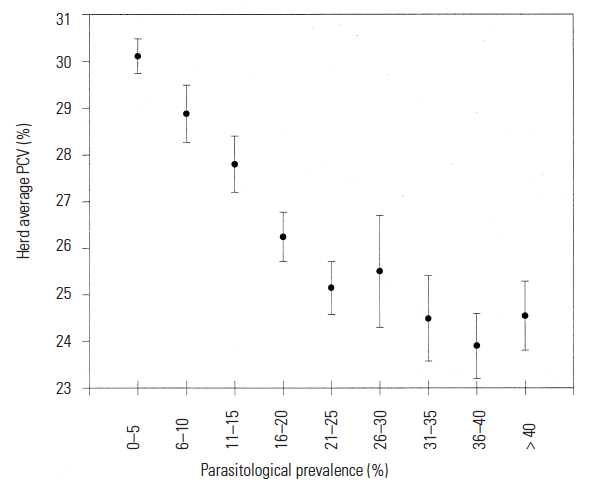
A recently described kit for in vitro isolation (KIVI) of trypanosomeshas proved promising for isolating and amplifying T. b. gambiense in humans, domestic and game animals.287 The test’s value in isolating T. congolense and T. vivax is still unknown. Since it is based on the cultivation of procyclic forms of trypanosomes, species differentiation is not possible.152
A miniature anion-exchange technique has been described for field use in the diagnosis of human trypanosomosis, 154, 171 but is too cumbersome for routine use in veterinary practice. The use of a minicentrifuge with buffycoat technique has been advocated.147 It is not satisfactory in bovine practice where large numbers of samples often have to be examined.
In the diagnosis of human trypanosomosis the cerebrospinal fluid (CSF) is examined for the presence of inflammatory cells, raised protein levels and parasites. For parasitology, the CSF is examined after centrifugation; the sediment obtained from 2 to 5 ml CSF is placed under a cover slip and examined microscopically for motile trypanosomes. 311
The non-specific diagnostic tests to detect raised protein levels in the serum of animals suspected to have trypanosomal infections148 have no role in diagnosis today. The development of anti-trypanosomal antibody detection techniques has been a major improvement in the serodiagnosis of trypanosomosis. The indirect immunofluorescent antibody test (IFAT)327 has been and still is used widely to diagnose trypanosomosis. The test has undergone several modifications so that it can differentiate, to a limited extent, between trypanosome species in ruminants.146 The serodiagnosis of trypanosomosis has greatly benefited from the introduction of enzyme immunoassays. The enzyme-linked immunosorbent assay (ELISA) was first used to detect antibodies against T. b. rhodesiense in humans.301 It was furtherdeveloped for use in animal trypanosomosis167 and was recently modified for large-scale use in trypanosomosis surveys.115 Antigens can be prepared using bloodstream forms or procyclic trypanosomes.101 Purified antigens are used preferably to ensure that tests are standardized and optical density cut-off values do not vary greatly between batches of antigen.
The ELISA compares favourably with the IFAT169 and has been found to give results that correlate with the local history of trypanocide usage.45 However, even if a trypanosomal infection has been cured, anti-trypanosomal antibodies persist for several months25 and antibody detection tests do not distinguish between current and past infections. They can only provide a presumptive diagnosis. Although the prevalence of anti-trypanosomal antibodies often increases with increasing prevalence of trypanosomal infections in a herd, antibody detection methods are not suitable for monitoring disease challenge in trypanosomosis endemic areas. Nevertheless, antibody detection tests, especially the antibody-detection ELISA, have been very useful tools for determining the distribution of trypanosomosis.292, 295 This is especially the case in areas where disease prevalence is low and where trypanocidal drugs are used frequently. Moreover, the prevalence of anti-trypanosomal antibodies is a sensitive indicator of the impact of tsetse control operations on disease challenge and for confirming the disease-free status of animals in areas from which tsetse has been eradicated.
Another alternative to the parasitological diagnosis of nagana is the use of assays to detect trypanosome-specific antigen, or species-specific or subspecies-specific DNA. An antigen detection ELISA for trypanosomosis has been described225, 226, 227 but field evaluations of the test have given inconsistent results. Additional work is needed to develop the test for routine diagnosis of trypanosomosis. A polymerase chain reaction (PCR) method has been developed for the diagnosis of infections with African trypanosomes in humans, animals and tsetse flies.92 Specific repetitive nuclear DNA sequences can be amplified for T. vivax and each of the three T. congolense subgroups.57, 201 A common primer set is available for detection of the three T. brucei subspecies. The test requires specialized equipment and highly trained personnel, consequently it is not suitable for use in most laboratories. Sample collection has been simplified by adapting the test using blood spotted on filter papers143 so that a large number of samples can be processed at one time. This makes the test potentially suitable for large-scale surveys. However, the cost of PCR analyses prohibits its routine use in veterinary investigation. The PCR technique’s ability to detect latent or mixed trypanosomal infections in different hosts renders it suitable for research into the complex relationships between trypanosomes and their vectors and hosts.245
Despite the development of more sensitive, more sophisticated and expensive diagnostic methods, the clinician will, for the foreseeable future, have to rely upon examination of blood smears, buffy coat preparations and findings at necropsy to confirm diagnoses of trypanosomosis. Accurate diagnosis is crucial to subsequent interventions to control nagana, and to monitor their success. At the herd level, the confidence placed in the results will largely depend on the size of the sample. In some cases, however, repeated sampling of an individual animal may be necessary before a diagnosis can be confirmed.
Differential diagnosis
In its various stages trypanosomosis resembles a number of other disease conditions, and it frequently occurs at the same time as other infections. In the acute febrile stage, trypanosomosis must be differentiated from redwater (babesiosis), anaplasmosis and East Coast fever. The haemorrhagic syndrome in acute T. vivax infection can be distinguished from anthrax and haemorrhagic septicaemia (Pasteurella multocida infection) by examination of Giemsa-stained blood smears, when large numbers of trypanosomes are found.
Acute trypanosomosis rarely produces icterus and is not accompanied by haemoglobinuria. These features and parasitological findings differentiate trypanosomosis from redwater. The two infections, however, may be concurrent. Similarly, anaplasmosis can occur in trypanosomeinfected cattle. On its own, anaplasmosis often produces icterus, as well as fever and anaemia; there may also be enlargement of lymph glands. Differentiation relies upon detection of the parasites in blood smears. Although lymph node enlargement and fever occur in East Coast fever, animals often cough and have diarrhoea. The presence of schizonts in stained smears of lymph node material and of piroplasms in blood smears confirms East Coast fever. At necropsy, the abomasal ulceration and kidney lesionscharacteristic of East Coast fever are not seen in trypanosomosis. Carcasses of animals which have succumbed to trypanosomosis are often oedematous and anaemic.
Chronic trypanosomosis is an afebrile disease, in which anaemia, emaciation and lymph node enlargement are prominent findings. It is important to differentiate this disease from malnutrition and helminthosis. In neither of these two conditions is lymph node enlargement found. The detection of helminth eggs in faeces is a useful adjunct to the diagnosis of helminthosis, but the absence of eggs in faecal samples taken from cattle with chronic fasciolosis, and the occurrence of anaemia and subcutaneous oedema can make differential diagnosis difficult. In cases of malnutrition, the degree of anaemia is rarely as extensive or severe on a herd basis as it is with trypanosomosis, although marked individual variation occurs. The late stage of enzootic bovine leukosis is characterized by weight loss and lymph node enlargement, but, unlike animals suffering from chronic trypanosomosis, appetite is diminished and anaemia is rarely severe.
When T. theileri is found in blood smears it is easily distinguished from other trypanosomes by its large size. Although T. theileri may occur in cattle infected with another species of trypanosome, the careful examination of thin Giemsa-stained blood smears is more likely to reveal the presence of mixed infections of tsetse-transmitted trypanosomes.
Clinical signs, pathology and diagnosis in goats and sheep
Trypanosomosis of goats and sheep has been stated to be of little significance because natural infections are seldom detected. 273 However, trypanosomosis of small ruminants is a major constraint on their health and productivity,102 and trypanosomal infections in goats and sheep are known to beprevalent in parts of southern Africa, occurring in Mozambique271 and Zambia.21 Small ruminants have a much shorter generation time than cattle, and it is likely that in tsetse areas this factor has produced greater natural selection for trypanotolerance in these species than has occurred with cattle. This may explain the fact that goats and sheep are found apparently thriving in tsetse-infested areas in which cattle cannot be kept without the use of drugs. Compared with exotic breeds, the local breeds of goats and sheep in the tsetse belts of East Africa are more tolerant of experimental trypanosomal infections,103 but trypanosomosis does nonetheless cause significant production losses even among local breeds.20, 21, 141
Of the salivarian trypanosomes, T. vivax is apparently the least pathogenic to these hosts, often producing mild infections. Trypanosoma congolense is an important pathogen and usually produces subacute or chronic disease whilst T. brucei infections may follow an acute, rapidly fatal course, especially in experimental infections. Frequently, however, naturally acquired T. brucei infections are chronic.273 Under natural conditions mixed trypanosomal infections are probably very common, although on parasitological examination one species usually predominates. Trypanosoma simiae also infects sheep and goats, but reports of clinical infections are rare.137, 172
Clinical signs
The general clinical signs of trypanosomosis of goats and sheep are similar to those in cattle. Acute, subacute and chronic disease syndromes occur naturally in goats and sheep in East Africa,103 and have been observed in goats in Zambia.21
Acute disease is characterized by intermittent and increasing dullness. During bouts of fever, sick animals walk stiffly, or stand with head held low, ears drooping and tail flaccid. Earlymorning rectal temperatures may exceed 41 °C in the febrile periods, when the pulse is rapid and the breathing shallow and fast. Anaemia develops rapidly and the PCV falls from 0,35 l/l to 0,18 l/l or less in two to four weeks, when mucous membranes become pale. Affected animals rapidly lose weight and condition, and may die within a month of the onset of detectable parasitaemia. Although corneal opacity occurs in experimental infections with T. vivax318 and T. brucei,122 this lesion is not common in natural cases.
The signs of subacute trypanosomosis are less marked, and the course of the infection resembles that in cattle. Anaemia develops quite rapidly, lymph nodes become enlarged (Figure 12.12) and, in goats, a marked jugular pulse can be seen. Although death occurs some 10 to 12 weeks after the onset of parasitaemia, the condition of many animals does stabilize and gradually improves. The improvement may be apparently complete; PCV, live weight and general condition may return to normal, and fever and parasitaemia abate. Other animals show slight improvement, but they remain anaemic, emaciated, weak and chronically affected. Chronic infections are very common and may developwithout marked clinical signs. Parasites are rarely found in the blood and animals are afebrile. Clinically, it is difficult to distinguish such animals from those with helminthosis. Furthermore, the immunosuppression induced by trypanosomosis in goats105 and sheep173 permits the establishment of large helminth burdens, which thus exacerbate the clinical signs in affected animals.104


As with cattle, trypanosomosis of small ruminants is a herd or flock disease which reduces productivity. Fertility is markedly reduced.121 The birth rate of goats in a tsetse-infested area of Zambia has been improved by regular chemoprophylaxis, even though few trypanosomal parasitaemiaswere detected in untreated control goats.21 Results from eastern Africa also indicate the importance of subclinical trypanosomosis on goat productivity.109
Pathology
The pathology of trypanosomosis in goats and sheep does not differ greatly from that in cattle.
Because of their smaller size and lower maintenance costs, small ruminants are popular experimental animals, and there are many reports on the pathology of experimental trypanosomal infections in these species. Recently, great attention has been focused on the effects of trypanosomosis on endocrine functions of these animals. Tsetse-transmitted T. congolense infections induce weight loss and irregular oestrous cycles in goats,161 whilst the inoculation of goats with bloodstream trypanosomes alters plasma levels of several hormones. Levels of thyroxine,219 testosterone,303 and plasma cortisol215 are depressed while plasma prostaglandin F2 levels rise during experimentally induced trypanosomosis in goats.218
Functional abnormalities have also been demonstrated experimentally: semen quality of infected rams is reduced1, 5 and ovarian dysfunction occurs in infected goats.216 Marked testicular lesions have been reported in experimental infections. In T. brucei-infected sheep, initial oedema of the scrotum was followed by a hardening of the scrotal wall and atrophy of the testes.120 Histologically, the most severe inflammatory reaction affected the tunica vaginalis and was characterized by a thick fibrinous exudate on the surfaces which led to adhesions and organization. Diffuse atrophy of the seminiferous tubules was associated with intertubular oedema, mononuclear cell infiltration and fibrosis. Gross testicular atrophy also occurred in goats infected with T. congolense, 139 while, histologically, degenerative lesions were evident. Seminiferous tubules were atrophied and their lumina were devoid of spermatids. Whilst these findings concur with field observations of reduced fertility, the same is not true for the severe central nervous system pathology of experimental infections of goats.319 The oedema and inflammation of the choroid plexus observed in experimental T. vivax infections may occur in natural infections, but theunder-diagnosis of trypanosomosis and the common practice of slaughtering sick animals generally precludes full evaluation of the pathology of natural infections.
Diagnosis
The diagnosis of trypanosomosis in small ruminants is no different from that already discussed for cattle, but it is much more difficult because of the larger proportion of subclinical infections. In these cases very low parasitaemias occur in the absence of obvious clinical signs. It is undoubtedly these factors which are responsible for the serious underestimation of trypanosomosis in small ruminants.
Differential diagnosis
The main disease entity which resembles trypanosomosis in goats and sheep is helminthosis, especially haemonchosis. Anaemia, ill-thrift, weight loss, submandibular oedema and high mortality rates are common to the two disease complexes, although diarrhoea sometimes accompanies helminthosis. Parasitological techniques have to be used for differential diagnosis. Frequently the two problems are concurrent, and the final diagnosis and treatment must depend upon clinical judgement.
Clinical signs, pathology and diagnosis in pigs
In many parts of southern Africa, indigenous breeds of domestic pigs survive in trypanosomosis-endemic areas. Pigs are refractory to infection with T. vivax, but are mildly affected by T. brucei, T. congolense and T. suis. In contrast, T. simiae is highly pathogenic to improved breeds of pigs, but local breeds seem to escape the ravages of this lethal trypanosome species. Of zoonotic significance is the role of the domestic pig in harbouring the human parasites T. brucei gambiense181 and probably T. brucei rhodesiense.11, 144 However, this latter association has not been demonstrated in southern Africa.
Trypanosomosis of pigs in Africa has been comprehensively reviewed,121, 274 and only an outline of the problem is presented here.
Clinical signs
Trypanosoma brucei and T. congolense infections in pigs generally produce only mild disease.237 Occasionally anaemia occurs, which is accompanied by loss of condition and progressive weakness leading to incoordination. Trypanosoma suis infections are of minor clinical importance and have only rarely been recorded; infections run a chronic course, but may kill young pigs in less than two months.
Trypanosoma simiae infections in pigs eclipse in importance all other trypanosomal infections in this host. This parasite may kill pigs after an incubation period of only four to six days, and infected pigsmaycollapse and die within 24 hours of the first signs of disease. The introduction of the parasite to a piggery is usually by an infected tsetse fly, after which spread is believed to be by mechanical transmission involving biting flies, often found in large numbers in pig pens.
The course of infection is so short that pigs die before losing condition, although some animals may live for as long as eight to ten days. Affected pigs are dull and inappetant or completely anorexic. They have a stiff, unsteady gait and eventually become prostrate. Dyspnoea is evident and cyanosis may be seen.
Some pigs froth at the mouth and there may be diarrhoea. Rectal temperatures reach 41 °C but the extremities are usually cold.
The course of the disease within a herd is dramatic. After one or two early deaths, several more occur within a week, and after a short time there may be only a few survivors.
Pathology
The necropsy findings vary considerably, and there are no pathognomonic changes. The blood is usually cyanotic, clots slowly, and chicken fat clots may be found in the heart. Haemorrhages affect the epi- and endocardium, kidneys and serosal surfaces. The spleen is usually enlarged and the pulp soft with a ‘strawberry jam’ appearance; there is excessive serous or sero-sanguineous fluid in the pericardial sac and pleural and peritoneal cavities; the trachea may be filled with froth when there is also pulmonary oedema. Thoracic and abdominal lymph nodes are usually enlarged and oedematous, and may be haemorrhagic.
Diagnosis and differential diagnosis
A history of sudden death in a number of pigs, combined with clinical signs as described, is suggestive of T. simiae in areas adjacent to tsetse infestations. Although sudden death may arise from African swine fever, anthrax, and other infectious and non-infectious diseases, the presence of fulminating parasitaemias, readily detected in Giemsa-stained blood smears, confirms the diagnosis.
Chronic trypanosomosis should be differentiated from helminthosis and malnutrition.
Clinical signs, pathology and diagnosis in horses and donkeys
With the advent of motorized transport and mechanized cultivation, the role of the horse has diminished in many parts of the world, including southern Africa. Furthermore, the peoples living in or near tsetse-infested habitats in this part of Africa have no tradition of keeping horses, but in some areas donkeys are important. Horses do not survive long in the presence of infected tsetse flies, whereas donkeys are more tolerant of tsetse-transmitted trypanosomes and frequently appear to thrive in lightly infested habitats. Nevertheless, T. brucei causes acute disease in donkeys, and is also a most serious pathogen of horses. Trypanosoma congolense produces a more chronic, wasting condition, whilst T. vivax, although usually producing chronic and more mild disease in equids,174 has been known to cause acute infection in the horse.56, 275
Clinical signs
The general clinical signs resemble those seen in ruminants. Horses with acute trypanosomosis are very dejected. Intermittent parasitaemia occurs and is accompanied by fever; rectal temperatures may reach 40,5 °C and there is tachycardia. Oedema of the lower limbs occurs early in the course of disease, as affected horses are less active than normal. Subcutaneous oedema then affects the ventral thorax and abdomen. Oedematous plaques may form on the flanks, but dependent oedema is more pronounced. Weakness sets in rapidly. Initially, mucous membranes may be congested and icteric but they become pale as the disease progresses. Ophthalmitis is not a constant finding; iritis may occur and up to 20 per cent of horses clinically affected by T. brucei may develop keratitis and corneal opacity.117 Although lymph nodes become enlarged, they are not easily palpable. The condition of the animal deteriorates rapidly and weakness may progress to paraplegia. Oedema becomes increasingly severe. Animals die within two to four weeks from the onset of clinical signs.
Chronic trypanosomosis commonly occurs in horses and donkeys. Animals lose condition and weight (Figure 12.13) and the coat becomes harsh and dry. The development of anaemia follows a course similar to that in cattle. Animals become extremely weak and show signs of ataxia, but usually continue to eat. Subcutaneous oedema, initially affecting the limbs and ventral abdomen, may extend to the sheath, scrotum, perineum and occasionally the head. The condition is exacerbated by strongylosis and is fatal, often after several months’ duration. In a survey in the Sebungwe area of Zimbabwe, a high proportion of donkeys was found to harbour T. brucei and T. congolense infections.34 Such mixed infections in horses give rise to more severe disease.
Pathology
The general pathology of equine trypanosomosis resembles that occurring in cattle. A very full account of the histopathology of experimental T. brucei infections in horses was given by McCully and Neitz.189
The histological lesions in the central nervous system of horses with experimental chronic T. brucei infections were characterized by a severe pleocytosis of the meninges and an extensive subpial gliosis. There was also segmented demyelination of optic tracts and some other areas of white matter as well as grey matter. Extensive perivascular cuffing with lymphocytes, plasmocytes, large mononuclear and Mott cells also occurred. Histological changes in the visceral and other organs were not uniform, but included reactive lymph nodes, interstitial myocarditis, haemosiderosis and focal endophlebitic lesions in veins of the testes.
Diagnosis and differential diagnosis
The diagnostic methods for equids are essentially the same as those for cattle. It is often necessary to repeatedly test those individuals suspected to be suffering from trypanosomosis in order to confirm diagnosis.
Acute disease must be distinguished from African horse sickness, anthrax and babesiosis, in which high fever,. sudden onset of signs, congestion of mucous membranes and petechiation occur. Similarly, in chronic trypanosomosis the oedema has to be differentiated from that occurring in African horse sickness, and the anaemia from that of equine infectious anaemia. Chronic babesiosis may also produce signs similar to chronic trypanosomosis. The signs of weight loss, emaciation and oedema caused by dourine (T. equiperdum infection), strongylosis, malnutrition and dental disorders should be differentiated from those caused by salivarian trypanosomes.
The signs of weight loss, emaciation and oedema caused by dourine (T. equiperdum infection), strongylosis, malnutrition and dental disorders should be differentiated from those caused by salivarian trypanosomes.
Clinical signs, pathology and diagnosis in dogs
Dogs are of greater cultural than economic importance in Africa. The typical village mongrel survives in many tsetse-infested areas with little or no attention from its owner. These are usually small animals, used for hunting and herding,and as watch-dogs. The exotic breeds, selected for similar purposes, quickly succumb to trypanosomosis when they are exposed to tsetse challenge. Exposure of susceptible breeds may also occur as they are transported through tsetse-infested areas, or as small numbers of infected tsetse are carried to otherwise tsetse-free areas by vehicles.
Dogs have been incriminated as reservoir hosts of T. brucei gambiense, the cause of West African human sleeping sickness, but such infections are subclinical in dogs.180. On the other hand, T. brucei brucei is the most pathogenic trypanosome in dogs, producing more acute disease than T. congolense, although strain differences occur, some strains being highly virulent. Dogs are refractory to infection with T. vivax
Clinical signs
The general clinical signs of trypanosomosis in dogs resemble those in livestock, but the course and severity of the disease are modified by the virulence of the parasite and the susceptibility of the host. In acute disease the dog suddenly becomes dejected and inappetant. Pulse rate, respiratory rate and rectal temperature are raised. Anaemia develops rapidly; mucous membranes become pale and the dog shows signs of weakness and depression. There is progressive weight loss; as the condition deteriorates, the coat becomes dry and subcutaneous oedema, particularly of the head and limbs, may develop.
Ocular lesions characterize both T. brucei and T. congolense infections but are more severe in the former case. Trypanosoma brucei invades all tissues of the eye, producing blepharitis, conjunctivitis, keratitis and uveitis. There may be transient glaucoma,199 and excessive lachrymation is common.
Additionally, T. brucei invades many other tissues, and by entering the cerebrospinal fluid causes ataxia and partial paralysis. Death ensues within 4 to 10 weeks of the onset of signs.
Infections with T. congolense generally follow a more protracted course. In chronic cases, dogs become severely emaciated, and ulceration of the oral and gastrointestinal mucosa may develop, leading to haemorrhage and melaena.110 Inappetance may only be intermittent, with animals being periodically ‘off colour’. In exotic breeds death occurs after only a few months, but indigenous dogs may remain infected for long periods.
Pathology
Lesions of trypanosomosis in dogs are broadly similar to those found in livestock. Anaemia is the most readily detectable change early in the course of infection, and the PCV drops markedly. Whilst T. congolense infections produce extensive interstitial oedema and ulceration of gastrointestinal mucosa, tissue lesions induced by T. brucei are much more severe. There is marked cellular infiltration and cellular degeneration and necrosis.199 The heart, choroid plexus and eyes are consistently and severely affected. There are also marked changes in lymph nodes due to the initial proliferation of lymphoid cells, although these may subsequently be depleted.
Diagnosis and differential diagnosis
Standard diagnostic methods are used to detect trypanosomes in the blood. However, after the initial phase of high parasitaemia, repeated sampling over several days may be necessary before trypanosomes can be found.
The anaemia caused by trypanosomosis must be differentiated from that arising from infection with Ancylostoma caninum, Babesia canis or Ehrlichia canis. Hookworm infection may be concurrent with trypanosomosis and can be confirmed by faecal examination. In young dogs, babesiosis (or biliary fever) is usually acute; rectal temperature may reach 41 °C and intraerythrocytic piroplasms are usually readily detectable in Giemsa-stained blood smears. The diagnosis of ehrlichiosis (Nairobi bleeding disease, tropical canine pancytopenia or tick fever) in an endemic area relies on the presence of such clinical signs as a moderate fever, crusty mucopurulent nasal and ocular discharges, foetid breath, stained teeth and pale mucous membranes.
Bilateral corneal opacity produced by trypanosomosis must be distinguished from that seen in canine hepatitis (adenovirus-2 infection), in which vomition and acute abdominal pain are presenting signs.
Control with trypanocides
Under light tsetse challenge a therapeutic approach to control can be successful and economical. Vigilance is necessary to ensure that infections are treated at an early stage, before disease has progressed too far and caused production losses. In these circumstances the need for supportive therapy is greatly reduced. However, the cost of constant monitoring has to be borne in mind. There are numerous well-documented examples of susceptible breeds of livestock being successfully reared in many tsetse-infested areas of Africa as a result of chemotherapy or chemoprophylaxis 113, 163, 182, 286 Considerable success in maintaining cattle, in the face of heavy trypanosome challenge, has been achieved in Zimbabwe30 and in Mozambique.279 To control trypanosomosis successfully by chemoprophylaxis, it is necessary to treat a high proportion of cattle at risk at regular intervals. This is not always possible, especially if owners have to pay for treatment, and consequently a therapeutic approach to control is widely used. There is considerable evidence that such therapeutic control assists the acquisition of protective immunity to locally prevalent trypanosome serodemes,113, 321 whereas with chemoprophylaxis immunity to trypanosomes does not seem to develop.114
Treatment
The satisfactory treatment of trypanosomosis requires more than a correctly administered trypanocidal drug, and the speed of recovery is largely determined by the plane of nutrition, the amount of exercise during convalescence and the duration of the disease. Well-rested and well-fed animals recover more rapidly after trypanocidal therapy than do undernourished animals which have to trek long distances to pasture and water. However, chronic trypanosomosis often fails to respond to therapy; the ferrokinetic disturbances and accompanying dyshaemopoiesis appear to be irreversible, and affected animals may remain thin and anaemic despite trypanocidal treatment.
History of trypanocidal drugs
Not all compounds active against trypanosomes are directly lethal to the parasites, and consequently these agents should be referred to as antitrypanosomal drugs. However, it is usual to refer to this category of medicines as trypanocidal drugs, or trypanocides. This latter term is used in the following account.
The small but important group of chemotherapeutic and chemoprophylactic compounds has been frequently reviewed.83, 157, 239, 325, 326 Trypanocides have a history which dates back to the early part of the twentieth century, when hundreds of substances were tested for antimicrobial activity and for activity against the then recently discovered trypanosomes, the first trypanocidal substances to be found being several dyes. Since then arsenical and antimonial compounds have been shown to be active against trypanosomes, and sodium antimony tartrate (better known as tartar emetic) became widely used throughout Africa, trypanosome-infected cattle receiving an intravenous course of the drug. Although this treatment was first used in Zimbabwe (then Southern Rhodesia) with great success, it was claimed that tartar emetic was used in Zambia (then Northern Rhodesia) on a scale unrivalled elsewhere.7
In 1921, suramin was marketed to treat T. evansi infections and human sleeping sickness. The drug is still widely used, nearly 80 years later, and is the drug of choice for the treatment of early cases of sleeping sickness. Research which began in the 1930s on phenanthridinium compounds led to the discovery of dimidium bromide. By the early 1950s this compound was used extensively both for therapeutic and for prophylactic purposes. Dimidium is relatively toxic; it produces reactions at the site of inoculation, and treated animals may become photosensitive. Despite this, heavy reliance was placed on the drug, and in the Lower River district of Malawi (then Nyasaland) cattle were successfully maintained as long as they received regular treatments of dimidium. However, in Malawi,188 as elsewhere, drug resistance to dimidium bromide emerged in the mid-1950s, and this drug was superseded by quinapyramine, which became available in the early 1950s. Serious drug resistance also emerged to this compound and by 1977 it had been withdrawn from the market. In 1955, homidium was marketed, but once again resistant strains of trypanosomes emerged and became widespread. At almost the same time, diminazene became available and was used to overcome quinapyramine- resistance. Being babesicidal as well as trypanocidal, diminazene has the advantage that it may also be used for treating animals suffering from redwater. Pyrithidium bromide then became available; it was marketed in 1956, but, again due to the emergence of drug resistance, it was withdrawn from the market in 1985.
When isometamidium was marketed in 1961 the active search for trypanocides diminished. This coincided with the period when many African countries gained independence from colonial rule and increasingly stringent requirements for the registration of drugs were demanded worldwide. Furthermore, comparable investments in pharmaceutical areas other than trypanocides are potentially much more profitable, so at the end of a century in which great scientific advances have been made, Africa remains with only four compounds (Table 12.2) with which to treat and prevent one of the continent’s biggest scourges.
Principles of trypanocide usage
The detailed pharmacokinetics and modes of action of the trypanocides are incompletely understood, but these compounds are regarded as having either therapeutic or prophylactic properties.149, 214, 240, 282 Diminazene and homidium are therapeutic trypanocides, although homidium does provide prophylaxis for several weeks.59 Isometamidium, whilst having a curative effect, is widely regarded as being the only trypanocide worthy of being used for prophylactic purposes. 150 The duration of chemoprophylaxis is influenced by several factors,232 which largely relate to the rate of excretion of the drugs.113 Diminazene is more rapidly excreted than isometamidium. After injection, initial high drug levels fall rapidly, although in cattle residual protection may last for up to 12 days after administration at a dosage rate of 7,0 mg/kg.309
The duration of protection conferred by isometamidium is dose-dependent,242 a higher dose providing longer protection. Experimentally, the prophylactic effect of isometamidium has been increased significantly by incorporating the drug in polymers in order to produce a sustained release device which is implanted subcutaneously.58, 89, 90 However, results from experiments with laboratory-adapted strains of parasites in relatively stress-free experimental animals, may bear little relation to events in the field. There are numerous reports that the higher the tsetse challenge, the shorter the duration of prophylaxis. Additionally, stressed animals on low planes of nutrition or with intercurrent disease do not always receive the same level of protection as do better managed animals. The most important factor determining the duration of prophylaxis is the susceptibility of the trypanosome population to the drug.241
Trypanocides are toxic compounds and have narrow therapeutic indices, so therefore the dosage rates cannot be greatly increased to obtain greater trypanocidal efficacy. Furthermore, they are not equally effective in the treatment of all species of trypanosomes. A single injection of diminazene cures T. congolense and T. vivax infections at a dosage rate of 3,5 mg/kg but a dosage rate of 7,0 mg/kg is necessary to treat T. brucei infections. The efficacy of the phenanthridine compounds, homidium and isometamidium against T. congolense and T. vivax is more marked than against T. brucei at recommended dosage rates. Dosage rates of isometamidium in excess of 2,0 mg/kg are toxic to cattle, and even below this level, the drug is irritant and has marked local effects.
The prophylactic effect of isometamidium depends upon the slow release of the drug from the depot created at the intramuscular injection site. Diminazene is rapidly excreted and has a negligible prophylactic effect. Homidium is absorbed more slowly than diminazene from an intramuscular injection site, but more rapidly than isometamidium. Isometamidium is irritant, and, at the site of injection, it induces an intense inflammatory response and causes necrosis of muscle fibres. This lesion becomes encapsulated and is later organized by fibrous connective tissue. From this depot, the drug is slowly released. Attempts to overcome this irritant side-effect by reformulation of the drug have been unsuccessful to date. Two important practical points are related to the injection site reaction. Firstly, the repeated inoculation of cattle kept under tsetse challenge induces extensive muscular scarring, and, to reduce carcass damage and subsequent losses from condemnation, the drug is usually administered into the less economically valuable neck muscles, often with untoward side-effects. Secondly, from the viewpoint of human consumption of treated animals, the question of drug residues in the animals’ tissues arises. It is thought that because of the poor absorption of the drug from the gastrointestinal tract,150 and the lack of untoward reports of toxicity in humans, the risks posed to human health by residues in meat and milk are minimal.
Table 12.2 Generic and trade names of trypanocides for the treatment of African animal trypanosomosis
| COMPOUND | MANUFACTURER | ACTION | RANGE OF DOSAGE RATES MG/KG | ROUTE OF ADMINISTRATION | REMARKS | |
|---|---|---|---|---|---|---|
| GENERIC NAME | TRADE NAME | |||||
| Diminazene aceturate | Berenil | Hoechst A.G., Germany | T | 3,5–7,0 | i/m | Also babesicidal. |
| s/c | Toxic to horses, donkeys, dogs and camels | |||||
| Homidium bromide | Ethidium | CAMCO Animal Health, U.K. | T(P) | 1,0 | i/m | |
| Homidium chloride | Novidium | Rhone-Merieux, France | T(P) | 1,0 | i/m | |
| Isometamidium | Samorin | Rhone-Merieux, France | P/T | 0,25–1,0 | i/m, (i/v)*** | Toxic above 2,0 mg/kg. Highly irritant. Avoid subcutaneous administration |
| Trypamidium | Rhone-Poulenc Sante, France | P/T | 0,25–1,0 | i/m, (i/v)*** | ||
| Quinapyramine sulphate* | Trypacide | Rhone-Merieux, France | T | 3,0–5,0 | s/c | Rest animals before and after treatment |
| Quinapyramine prosalt | Trypacide prosalt** | Rhone-Merieux, France | P/T | 3,0–5,0 | s/c | Dosage calculated as sulphate |
* Reintroduced in 1985 to treat mainly T. evansi infections
** Prosalt is a mixture of sulphate and chloride salts of quinapyramine
*** Source: reference 63
i/m = intramuscular
s/c = subcutaneous
i/v = intravenous
T = Therapeutic action
P = Prophylactic action
(P) = Short prophylactic activity
Trypanocide use in cattle, goats and sheep
These animals tolerate diminazene, homidium and isometamidium at recommended dosage rates (Table 12.2), but quinapyramine is no longer used in these species. Trypanocides are dissolved in water immediately before use, and normal aseptic precautions are necessary in order to reduce reactions at the injection site. Work oxen and dairy animals should receive deep intramuscular injections into the gluteal muscles instead of the neck muscles. Calves, goats and sheep receive deep intramuscular injections in the thigh, care being taken to avoid injury to the sciatic nerve. The intravenous administration ofisometamidium as a 1 per cent w/v solution has been successfully employed to treat bovine trypanosomosis,63 but careful injection is essential to avoid systemic and local reactions.15
The practice of using different trypanocides alternately to overcome drug resistance warrants mention. Diminazene should be administered not less than one month after isometamidium treatment,9 and should be followed by an interval of at least one week before further isometamidium treatment to avoid lethal synergistic toxic effects when normal dosage rates of the two drugs are administered a short time apart. Affected cattle lose condition rapidly, becoming dehydrated and emaciated. Animals may recover after a long time, but up to 6 per cent die. Necropsy findings usually include severe fatty changes in the liver, with enteritis being more variable.
Trypanocide use in pigs
Chemotherapy of T. simiae has limited success because by the time signs of disease are seen it is usually too late to save the animal’s life by treatment.
Even when early cases are treated, few drugs are effective. Simultaneous treatment with quinapyramine sulphate at 5,0 mg/kg and diminazene at 10,0 mg/kg can effect a cure. Other dosage rates of these drugs have also been used, but relapses after treatment are common and may be due to the presence of parasites in the cerebrospinal fluid.334 Constant vigilance and retreatment are therefore necessary. In the face of an outbreak it is usual to treat all pigs at risk prophylactically in order to halt the spread of infection, and for this purpose quinapyramine prosalt has been used at a dosage rate of 11,7 mg/kg (calculated as the sulphate) with variable success. Isometamidium given as a 4 per cent w/v solution by deep intramuscular injection at dosage rates of between 15,0 and 25,0 mg/kg are effective in halting the spread of disease, but large, crater-like ulcers develop at the sites of injection.84
Prophylaxis has been attempted with a suraminquinapyramine complex at 40 mg/kg and with quinapyramine chloride at 50 mg/kg, but neither of these formulations is commercially available.
Trypanocide use in equine infections
Diminazene is poorly tolerated by donkeys31 and, on occasion, horses. The use of the higher dosage rate (7,0 mg/kg) required to treat T. brucei infections is often accompanied by nervous signs. Isometamidium may be used to treat and prevent equine T. congolense and T. vivax infections, but careful aseptic precautions and attention to the intramuscular injection technique are needed to reduce local tissue reactions. Separate needles should be used for filling the syringe and giving the injection. To avoid depositing the irritant solution subcutaneously, a sufficiently long needleshould be used; leak-back should be reduced by using a small-gauge needle, applying firm pressure as the needle is withdrawn and massaging the injection site well to disperse the solution in the muscle. Quinapyramine sulphate may be used to treat T. brucei, T. congolense and T. vivax infections, but animals must be well rested beforehand, since side-effects usually occur, which usually pass within one to two hours. These are transient and include restlessness, salivation, sweating, fasciculations, diarrhoea and abdominal discomfort, which may cause the animal to back away from the handler or to become recumbent and roll. In weakened animals the dose should be divided in two; the second half should be administered by subcutaneous injection four to six hours after the first.
Horses and donkeys can be protected by quinapyramine prosalt given at a dosage rate of 3,0 mg/kg. Since the prosalt is insoluble, it is administered as a suspension by subcutaneous injection. The injection site should be massaged well to minimize the local reaction, but a depot of drug is formed in the tissues at the injection site, which induces an inflammatory and fibroblastic response. The nodule may ulcerate and lead to discharge of the drug to the exterior.
Trypanocide use in dogs
Diminazene is toxic to dogs at doses higher than 3,5 mg/kg and its use is best avoided in the treatment of trypanosomosis, especially of T. brucei infections which require treatment at 7,0 mg/kg. However, simultaneous administration of difluoromethylornithrine (DFMO) and diminazene has resulted in the successful treatment of T. brucei infections.6 Homidium, at a dosage rate of 1,0 mg/kg, or isometamidium, at a dosage rate of between 0,5 mg/kg and 1,0 mg/ kg, may be used to treat T. congolense infections. However, the intramuscular injection of these drugs often produces hot, painful inflammatory lesions at the injection site, which results in lameness of varying severity. Treated dogs should be well rested until the inflammation has subsided.
Quinapyramine sulphate has been found to be suitable for the protection of dogs against trypanosomosis,10 and can also be used for treatment. Animals must be cool and rested before and for at least two hours after subcutaneous injection of quinapyramine sulphate; a dosage rate of 5,0 mg/kg is used, and may be repeated at monthly intervals for prophylaxis.
Failure of trypanocides
When trypanocides fail to provide the expected cure or protection there is a tendency to assume that drug resistance has arisen. Whilst this may be true, there are several other reasons which can contribute to their failure (Table 12.3). Considerable logistical constraints exist throughout Africa in the provision of veterinary services to rural areas, and farmers often resort to the treatment of their own animals. A survey conducted in the Eastern Province of Zambia, for example, revealed that 67 per cent of the trypanocides were administered by the cattle owners.61 Trypanocides are relatively inexpensive drugs. They are sold in granular, tabletted or powder form and have to be dissolved in water to obtain a solution for injection. Factors which contribute to their widespread underdosage are incorrect dilution and the inaccurate estimation of the live weights of animals. The use of unsterilized syringes, needles and the water in which the drugs are administered may induce the formation of abscesses which, if they burst to the exterior, results in the loss of the drug. Consequently, treatment quite often fails to produce the expected results.
The use of trypanocides at subtherapeutic levels is believed to promote drug resistance in trypanosomes.89, 322 Since diminazene is rapidly excreted, the risk of trypanosomes being exposed to sublethal levels of the drug is less than is the case with isometamidium. Prophylactic levels of isometamidium wane over much longer periods. Even when cattle receive isometamidium at the correct dosage rate, it may be administered at the wrong time. Tsetse challenge may increase at a time when drug levels have declined and are not protective. Furthermore, it appears that the duration of the protection is reduced in those cattle which are in poor general health or are on a low plane of nutrition, and it is thought likely that trypanocides require an active immune response by the host for them to be fully effective.113
Table 12.3 Causes of apparent drug resistance in trypanosomes: failure of trypanocides
| Underdosage | |
| underestimated body weight overdiluted solution of trypanocide incorrectly calculated dose volume deliberate underdosage to enable treatment of more animals incorrect injection technique
| |
| Incorrect strategy | |
| irregular treatment prolonged intervals between treatments (challenge when drug levels have waned) lack of knowledge of seasonality | |
| Stress-induced metabolic changes | |
| malnutrition lactation intercurrent disease breed-associated stress work and trekking | |
| Increase in challenge | |
| seasonal changed grazing area absolute increase in tsetse numbers | |
| Relapsing infections | |
| True drug resistance | |
A serious cause for the failure of trypanocides is the phenomenon of relapsing infections.113 Trypanosomes in extravascular sites may not be exposed to lethal concentrations of trypanocides, and they may then emerge at a time when drug levels in the blood have declined. Trypanosoma brucei in the cerebrospinal fluid is not exposed to trypanocidal levels of diminazene, which does not readily cross the blood–brain barrier.317 Although trypanosomes are cleared from the blood after treatment, the infection may relapse as parasites reinvade the bloodstream from druginaccessible sites. A similar event occurs with T. vivax178 which is known to invade the aqueous humour of the eye and the cerebrospinal fluid of goats.318 The extent of the occurrence of relapsing infections under natural conditions is not known, as it is difficult to distinguish between true drug resistance and a relapse in the field, and the occurrence of a relapsed infection may often simply be ascribed to ‘resistance’.
True resistance to trypanocides is a spectre which looms large. The development of resistance has led to the withdrawal of many trypanocides in the past, and resistance to the few remaining currently used trypanocides is documented.89, 239 The degree of resistance in trypanosomes is not clear cut; a spectrum of drug sensitivity occurs, and resistance to low levels of a drug may be overcome by increasing the dosage rate.113 Cross-resistance occur.325 Multiple resistance of a strain of T. vivax to normal therapeutic dosage rates of isometamidium, diminazene, quinapyramine and homidium has been confirmed, 258 but fortunately such cross-resistance is rare. Resistance to quinapyramine developed after only six months in some areas of Kenya321 — a sanative treatment of either homidium or diminazene was administered to control this. Cattle thus treated and removed from the challenge area were subsequently found to be free of infection and also grew well.321 Nevertheless, induction of resistance to quinapyramine in T. congolense has been shown to result in cross-resistance to homidium, isometamidium and diminazene.229 Hence, quinapyramine should not be used in cattle. Diminazene and isometamidium are regarded as a sanative pair, since cross-resistance between the two is very rarely seen in the field113 and, in fact, the multiple resistance in T. vivax mentioned above258 was overcome with diminazene at 7,0 mg/kg.
In southern Africa, the only well-substantiated report of resistance to isometamidium was made from Zimbabwe in 1974.160 Infection of two non-immune cattle with the suspected isometamidium-resistant T. congolense strain was not cured by isometamidium treatment at l,0 mg/kg, although treatment at 2,0 mg/kg did effect a cure. The problem in the field was overcome by herd sanative treatments with diminazene. More recently, diminazene aceturate resistant stocks of T. congolense have been isolated from cattle in the Zambezi Valley of Zimbabwe138 and diminazene and isometamidium resistance has been reported in Zambia.37 In cases of serious drug resistance, cattle have to be withdrawn from the tsetse area, but resistant trypanosomes are cyclically transmitted by tsetse, and they may then enter the wild host reservoir. Transmission of the strain will then continue, albeit much diluted, between wild hosts.99, 100, 134 The reintroduction of livestock into such areas may quickly lead to the re-emergence of drug resistance.
Methods to prove easily that certain strains of parasites are resistant are not available. Currently, three techniques are commonly used to detect drug resistance. Firstly, tests in ruminants provide direct and accurate information on the dose required to cure a trypanosomal infection.269 The frequently used mouse inoculation tests can, at best, only broadly indicate the sensitivity of a strain.269 Secondly, attempts to conduct in vitro bioassays with culture-adapted trypanosomes are being made, but not all field strains will adapt to culture systems and, of those that do, some may acquire an altered pattern of drug sensitivity.140 Thirdly, as an alternative to bioassays for determining the sensitivity of trypanosome strains, is the use of a trypanocidal drug-ELISA in conjunction with parasite detection tests. A competitive ELISA capable of detecting small amounts of isometamidium in the serum of cattle has been developed74, 75, 319 and has been validated under field conditions.73
Strategic use of trypanocides
The single rainy season in southern Africa influences tsetse dispersal and leads to a seasonal peak in the incidence of trypanosomosis. 44 The determination of the precise trypanosome challenge is a complicated matter,255 but from the practical point of view it is more important to establish the disease incidence or risk. This tends to be seasonal, with many new infections being contracted soon after the beginning of the rainy season. This is the time at which isometamidium chemoprophylaxis should be used. Repeated treatments may be needed during the rainy season, and cattle can be maintained by chemotherapy with diminazene during the lower risk period. Evidence pointing to the potential value of this strategy has been obtained with sentinel cattle in southern Tanzania, which also has unimodal rainfall.47
To apply this or any other strategy it is necessary to meet all of the logistical requirements of drug administration and disease surveillance. It is frequently the logistical bottlenecks which reduce the efficacy of trypanocides rather than the inherent inactivity of the drugs.113 The cattle owner’s attitude towards his animals and the management of disease in his animals is an important factor contributing to the success or failure of drug use. A survey of trypanocide use in several countries of southern Africa61 showed that, despite the availability of diminazene and isometamidium, cattle owners preferred to treat animals that were clinically affected using a curative rather than a prophylactic approach. Moreover, preference was given to the treatment of the more productive animals, such as oxen and cows. Although this treatment regime does not improve the reproductive performance of cows, it significantly reduces mortality due to nagana and improves the condition of oxen. This may, from a cattle owner’s point of view, be the most effective way of dealing with trypanosomosis. It is not evident that other strategies will be readily accepted.
The warning has been sounded: ‘Cattle cannot be kept alive indefinitely on the end of a needle’,176 and this must be recognized at the outset of any control programme. The use of trypanocides can be highly effective when there is close veterinary supervision, but it is only palliative. Attempts must be made to reduce tsetse numbers and trypanosome challenge. In the short term, the benefits which accrue from the control of trypanosomosis with trypanocides amply justify the costs. In the long term this approach is not sustainable.
Trypanosomosis control strategies
For more than 60 years, governments in southern Africa have made concerted efforts to control tsetse in their battle against trypanosomosis. The ultimate goal, and the basis of many control strategies, was the eradication of the tsetse fly. The Tsetse and Trypanosomiasis Control Branch of Zimbabwe’s Department of Veterinary Services has achieved notable success in controlling the disease by progressively removing the vector. Different strategies have been used, and within each strategy different methods have been applied as they have become available. A brief consideration of trypanosomosis control in Zimbabwe is thus very instructive.
Vector control began in the 1930s when wild hosts of tsetse were eliminated by large-scale hunting operations. This method of control was abandoned in 1960. Tsetse control then relied initially on bush clearing and the application of residual insecticides to tsetse resting sites by ground-based spraying teams. However, these methods on their own did not result in satisfactory control of nagana, and in 1964 selective hunting began in fenced corridors that delimited tsetse-infested regions. To limit the extent of the disease, livestock were excluded from many tsetse areas by well-maintained cattle fences. As a result of progressive operations, tsetse were well under control by the early 1970s, and a system of compulsory monthly inspection of all livestock, with the treatment of sick animals, effectively contained the disease. These gains were reversed during the liberation war years of the 1970s. In the mid-1970s an estimated half a million cattle were at risk of trypanosomosis. Every year some 140 000 blood smears were taken from cattle in risk areas, and approximately 100 000 treatments were given annually.44 From 1980, largescale aerial spraying of insecticides was used, which was aimed at reclaiming areas invaded by tsetse during the war. By 1989, trypanosomosis was again well under control. In the year to September 1989, two million cattle were inspected and, of 67 563 blood samples examined, only 596 cases were detected. From 1991 onwards, odour-baited, insecticide-treated cloth targets became the method of choice of the Zimbabwean Tsetse and Trypanosomiasis Control Branch and almost completely replaced aerial and ground spraying. By 1999, approximately 60 000 targets were deployed as an effective barrier against tsetse reinvasion from neighbouring countries and the tsetse-infested areas in the north of the country. The barrier protects more than 1 000 000 head of cattle, mainly in communal lands, from the threat of tsetse.62
Despite considerable gains in other countries, few achievements have been sustained because of the prohibitive cost of preventing tsetse from reinvading previously cleared areas in the absence of natural barriers. Nagana, therefore, remains a serious constraint to development throughout much of Africa and new approaches towards its control have been devised.
The new control strategy aims at intervening in areas where achievements are likely to be sustained by the beneficiaries. Consequently, in most cases, trypanosomosis will be controlled on a smaller, localized scale in areas that have to be selected carefully. Socio-economic and technical criteria are the major determinants in the selection process.
Control interventions are more likely to be sustained where the beneficiaries perceive trypanosomosis as a serious constraint to their development and where its control will result in significant economic gains. To identify such areas, accurate information is required on the distribution and socio-economic importance of the disease, and on farmers’ perceptions of the impact of the disease and its control.
The impact of nagana on rural development is often unknown and control operations have frequently been planned and implemented based on spurious or incomplete information and invalid assumptions. The socio-economic impact of bovine trypanosomosis varies widely and is influenced by factors such as the disease challenge, the degree of tolerance to infection and the application of control measures.
The long-term success of an intervention to control nagana (and many other diseases) does not depend entirely on its effect on livestock production. The livestock owner’s perception of its impact is of utmost importance. This perception will vary from area to area depending on the farming or livestock management practices and may vary in time, depending on other factors constraining development. For example, in areas where cattle are mainly used for draught power, the effect of trypanosomosis on fertility is likely to be perceived as a minor problem. In such areas, cattle owners are unlikely to sustain control measures that greatly improve reproductive performance. The opposite may be true in areas where the sale or accumulation of cattle is of prime importance. Even if nagana is perceived as a constraint, control interventions will not always result in financial and economic gains. This will normally be the case when either the potential for herd expansion is limited or where there is limited opportunity to increase the area under cultivation. These considerations apply, for example, to many parts of Malawi. Under such conditions, interventions to improve agricultural and livestock production may have minimal impact and are unlikely to be sustained by the livestock owner.
Several methods are available to control trypanosomosis. They may be directed against the vector or the parasite, or towards the livestock and modified management. In choosing a method, technical and economic aspects have to be considered and the most viable option selected.
Some of the tsetse control techniques that have been developed with large-scale eradication in mind are not well suited for use in localized, small-scale control operations. Currently, tsetse control utilizes odour-baited, insecticide-treated cloth targets and insecticide-treatments of cattle (see Vectors: Tsetse flies). Odour-baited, insecticide-treated targets have been very effective in Zimbabwe and have been employed in other African countries to control various species of tsetse under different ecological conditions. The concept of using insecticide-treated cattle to control tsetse has been revived since the development of synthetic pyrethroids for fly and tick control.
This method has been used successfully and, at first sight, its simplicity and its effect on ticks and biting flies have prompted demands for its more widespread use wherever cattle are exposed to tsetse.38, 60, 192 However, caution is necessary for several important reasons. The first relates to cost since pyrethroids are relatively expensive. For a commercial farm, changing from its usual acaricide to a pyrethroid could increase costs significantly. In areas where traditional farming methods are practised, tick control measures are not always routinely used, and the regular use of pyrethroids to treat cattle would be a high additional expense. In view of the high operational costs, the duration of treatment should be kept to a minimum. Therefore, before embarking on a control operation with this method, there should be evidence that it will be effective. Several important questions have to be asked: Do tsetse in the area take a high proportion of feeds from cattle? Can tsetse reinvasion of the area be successfully controlled? 304 How many animals should be treated and at what treatment interval? For how long must this measure be enforced? To continue to treat cattle with deltamethrin indefinitely to combat constant tsetse reinvasion raises not only the question of its cost, but also of the effectiveness of its insecticidal and acaricidal properties. The emergence of ticks that are resistant to deltamethrin would be serious, although this may take several years to develop.
The control of ticks in hitherto untreated herds may lead to instability in the relationships between cattle and tick-borne disease agents. The endemic stability prevalent in many traditionally owned herds (see Bovine babesiosis) is likely to be disturbed by rigorous pyrethroid treatment.294 To prevent losses, vigilant veterinary staff would need to be available to treat sick animals, and there are implications for the vaccination of cattle against tickborne diseases, which may rely upon moderate sustained challenge for continued immunity. Equally, with the cessation of treatment and increasing tick burdens, the intensity of transmission of Anaplasma spp., Babesia spp., Theileria spp. and Ehrlichia (Cowdria) ruminantium might then increase, leading to increased mortality rates. Despite these negative considerations, the method has much to commend it. It can have a rapid impact on tsetse numbers, but it should be used for the shortest time possible, in conjunction with other control measures. The management of such insecticide-treated cattle in an active, offensive way is also possible. Animals could be treated and moved deliberately into a tsetse focus in an attempt to reduce fly numbers quickly. Protection of these cattle against trypanosomosis would be necessary, because tsetse could still feed and transmit trypanosomes before being killed.293
Despite the proven effectiveness of insecticide-treated cattle and odour-baited targets, the success of a tsetse control operation will depend largely on its planning and implementation. Government-controlled interventions are normally carefully planned and implemented by highly skilled technical personnel.
Although community-driven tsetse control operations may still benefit from skilled personnel in the planning phase, the responsibility for the longterm implementation is likely to be in the hands of the beneficiaries. The degree of co-ordination and synchronization of activities between the community members will determine the effectiveness of such tsetse control operations. Synchronizing and co-ordinating the tsetse control activities of a heterogeneous group of community members, with diverse objectives, may be difficult. Therefore, where tsetse eradication cannot be achieved it is unlikely that tsetse control measures will be the only tool to control nagana, and a combination of methods is likely to be used. Depending on the perception of the disease impact, preference will usually be given to a particular method. Where endemic human sleeping sickness occurs, such as in parts of Uganda, there is evidence that communities will sustain the use of traps to control tsetse numbers.
Trypanocidal drugs will continue to play an important role in the integrated control of trypanosomosis. They appeal to communal cattle owners because they provide a means of protecting private goods andmayachieve impressive results at low costs. Although the low-level usage of these drugs may reduce mortality rates, such usage generally has low impact on animal production. This will be the case particularly in high challenge areas and when farmers use trypanocides therapeutically rather than prophylactically. The development of resistance in trypanosomes to trypanocides is a continuous threat to their sustainable use in the control of nagana. Even in areas where resistance to trypanocides has not yet been demonstrated, the probability of its development should influence the selection of an appropriate control strategy.
Millions of domestic animals are kept in tsetse-infested areas, but the degree to which they are exposed to tsetse is frequently determined by management practices. The avoidance of heavily infested watering points or grazing reserves reduces challenge, and thus controls trypanosomosis to some extent, but the availability of alternative water sources and supplies of feed are then necessary. Prevention of livestock movement into tsetse habitats also limits the disease, but the enforcement of livestock movement control in Africa is not easy, because of communal grazing practices, seasonal migrations and local political pressures usurping the law.
If animals are exposed to tsetse fly, the severity of the subsequent trypanosomal infections is modified by the animals’ general well-being. Well-fed animals are better able to resist the effects of trypanosomal infections, and many animals attain an equilibrium with the parasites. The provision of supplementary feed, especially during the dry season, is an important factor in mitigating disease.
Lactating animals, draught oxen and selected breeding bulls benefit from a high plane of nutrition; they are then better able to withstand the effects of trypanosomosis. The general principles of good management also require the prevention and treatment of other diseases, and regular, tactical tick control, anthelmintic treatment and vaccination are useful adjuncts to trypanosomosis control. Sadly, it is often the case that many livestock owners prefer a simple cure to the investment of their time, energy and resources in improved management. The role of good husbandry in disease control is frequently underestimated. Improved breeds of livestock not adapted to tropical conditions generally fare poorly in hot climates, where pastures are often of low quality and tick challenge is frequently encountered. They also readily succumb to trypanosomosis.
Immunological control
The antigenic complexity of trypanosomes has thwarted attempts to develop a vaccine.207, 222 Although potential immunological targets within the parasite have been identified, no vaccine will be commercially available in the near future, and the greatest hope for the immunological control of animal trypanosomosis lies in the exploitation of trypanotolerant breeds of livestock.213 Trypanotolerance is a general breed characteristic and, as such, some individuals are more tolerant than others.253 Furthermore, the trait is complex; it is related to the ability to resist anaemia, the ability to control parasitaemia and, possibly, the ability to mount a more effective immune response to the trypanosome.213 It also appears that while these processes are under genetic control, they are not directly linked to each other. The search for genetic markers has concentrated on the major histocompatibilitycomplex and on leukocyte antigens.285
In the meantime, the correlation of packed cell volume and productivity parameters of N’Dama cattle284 forms the basis of a field test to select young trypanotolerant animals.283 It is intended, through a process of selective breeding of trypanotolerant animals, to reduce the problem of trypanosomosis and to improve livestock productivity. However, this approach offers little hope for southern Africa today: inheritance is complex and the number of trypanotolerant animals is relatively small.213 On the other hand, the fact that some Zebu cattle survive in tsetse-infested areas provides a basis for attempting to select the more tolerant of these animals for breeding purposes. To do so would hasten the process of natural selection that is already occurring.
References
- AGU, W.E., IGE, K. & OLATUNDE, D.S., 1986. Evaluation of semen quality of rams infected with Trypanosoma vivax. Animal Reproduction Science, 11, 123–127.
- AKOL, G.W.O., AUTHIE, E., PINDER, M., MOLOO, S.K., ROELANTS, G.E. & MURRAY, M., 1986. Susceptibility and immune responses of Zebu and taurine cattle of West Africa to infection with Trypanosoma congolense transmitted by Glossina morsitans centralis. Veterinary Immunology and Immunopathology, 11, 361–373.
- AKOL, G.W.O. & MURRAY, M., 1982. Early events following challenge of cattle with tsetse infected with Trypanosoma congolense: Development of the local skin reaction. The Veterinary Record, 110, 295–302.
- AKOL, G.W.O. & MURRAY, M., 1986. Parasite kinetics and immune response in efferent prefemoral lymph draining skin reactions induced by tsetse-transmitted Trypanosoma congolense. Veterinary Parasitology, 19, 281–293.
- AKPAVIE, S.O., IKEDE, B.O. & EGBUNIKE, G.N., 1987. Ejaculate characteristics of sheep infected with Trypanosoma brucei and T. vivax: Changes caused by treatment with diminazene aceturate. Research in Veterinary Science, 42, 1–6.
- ANENE, B.M., ANIKA, S.M., AND CHUKWU, C.C., 1997. Effects of difluoromethylornithine after intravenous administration and its combination with diminazene aceturate against Trypanosoma brucei in experimentally infected dogs in Nigeria. Revue d’Elevage et de Médecine Vtérinaire des Pays tropicaux, 50, 221–225.
- ANON., 1929. Annual Report of the Veterinary Department of Northern Rhodesia 1928. Lusaka: Government Printer.
- ANON., 1959. Annual Report of the Department of Veterinary Services and Animal Husbandry for the Year 1959. Blantyre: Government Printer.
- ANON., 1967. Annual Report of the Branch of Tsetse and Trypanosomiasis Control, Veterinary Services, Ministry of Agriculture, Rhodesia, for the year ending 30th September 1967. Harare: Government Printer.
- ANON., 1971. Annual Report of the Branch of Tsetse and Trypanosomiasis Control, Department of Veterinary Services, Ministry of Agriculture, Rhodesia, for the year ended 30th September 1971. Harare: Government Printer.
- ANON., 1989. Tsetse Research Laboratory Annual Report 1989. Bristol: Tsetse Research Laboratory, Overseas Development Administration and University of Bristol.
- ASKONAS, B.A., 1984. Interference in general immune function by parasite infections: African trypanosomiasis as a model system. Parasitology, 88, 633–638.
- BACHMANN, M., 1990. Tsetse/nagana situation, Zululand. A provisional evaluation of nagana in Zululand. Report to the Director, Department of Animal Health. Pietermaritzburg: Department of Agricultural Economics and Marketing, Republic of South Africa.
- BAILEY, J.W. & SMITH, D.H., 1992. The use of the acridine orange QBC technique in the diagnosis of African trypanosomosis. Transactions of the Royal Society of Tropical Medicine and Hygiene, 86, 630.
- BALIS, J., 1977. Note sur la toxicité de l’Isometamidium par injection intraveineuse chez quelques mamifères et spécialement chez le dromedaire. Revue d’Elevage et de Médecine Vtérinaire des Pays tropicaux, 30, 373–375.
- BARRY, J.D. & EMERY, D.L., 1984. Parasite development and host responses during the establishment of Trypanosoma brucei infection transmitted by tsetse fly. Parasitology, 88, 67–84.
- BATEMAN, A., SINGH, A., KRAL, T. & SOLOMON, S., 1989. The immune-hypothalamic-pituitary-adrenal axis. Endocrine Reviews, 10, 92–112.
- BAYLIS, M., 1996. Effect of defensive behaviour by cattle on the feeding success and nutritional state of the tsetse fly, Glossina pallidipes (Diptera: Glossinidae). Bulletin of Entomological Research, 86, 329–336.
- BAYLIS, M. & NAMBIRO, C.O., 1993. The effect of cattle infection by Trypanosoma congolense on the attraction, and feeding success, of the tsetse fly Glossina pallidipes. Parasitology, 106, 357–361.
- BEALBY, K.A. & CONNOR, R.J., 1990. Department of Veterinary and Tsetse Control Services, Chipata, Zambia, and Regional Tsetse and Trypanosomiasis Control Programme, Harare, Zimbabwe. Unpublished observations/data.
- BEALBY, K.A., CONNOR, R.J. & ROWLANDS, G.J., 1996. Trypanosomosis in goats in Zambia. International Livestock Research Institute, Nairobi, Kenya.
- BERNARDS, A., 1984. Antigenic variation of trypanosomes. Biochemica et Biophysica Acta, 824, 1–15.
- BEUTLER, B. & CERAMI, A., 1988. The common mediator of shock, cachexia, and tumor necrosis. In: Advances in Immunology. London: Academic Press.
- BLACK, S.J., 1986. Host responses which control Trypanosma brucei parasites. In: MORRISON, W.I., (ed.). The Ruminant Immune System in Health and Disease. Cambridge: Cambridge University Press.
- BOCQUENTIN, R., VERY, P. & DUVALLET, G., 1990. Cinétique des anticorps après traitement trypanocide chez des bovins infecté expérimantalement ou naturellement. Intérêt épidémiologique. Revue d’Elevage et de Médecine Vétérinaire des Pays tropicaux, 43, 479–483.
- BOREHAM, P.F.L., 1979. Tsetse feeding patterns and the transmission of trypanosomes. Transactions of the Royal Society of Tropical Medicine and Hygiene, 73, 130–131.
- BOSE, R., FRIEDHOFF, K.T., OLBRICH, S., BOSCHER, G. & DOMEYER, I., 1987. Transmission of Trypanosoma theileri to cattle by Tabanidae. Parasitology Research, 73, 421–424.
- BOSMAN, P.P., 1990. Trypanosomosis (T. congolense) in South Africa. Office International des Epizooties: Disease Information, 3, 79–80.
- BOYT, W.P., 1978. Food hosts of tsetse. The Rhodesia Science News, 12, 141–143.
- BOYT, W.P., 1979. Trypanosomiasis in Zimbabwe Rhodesia. Rhodesian Veterinary Journal, 10, 54–63.
- BOYT, W.P., LAWRENCE, J.A., MACKENZIE, P.K.I. & FRENCH, R.L., 1971. The toxicity of diminazine in donkeys. Rhodesian Veterinary Journal, 1, 75–78.
- BOYT, W.P., LOVEMORE, D.F., PILSON, R.D. & SMITH, I.D., 1963. A preliminary report on the maintenance of cattle by various drugs in a mixed G. morsitans and G. pallidipes fly belt. Proceedings of the Ninth Meeting of the International Scientific Council for Trypanosomiasis Research, Conakry, 1962. Commission for Technical Cooperation in Africa. Hertford: Stephen Austin and Sons Ltd.
- BOYT, W.P. & MACKENZIE, P.K.I., 1970. A preliminary note on a virulent strain of Trypanosoma vivax in Rhodesia. Rhodesian Veterinary Journal, 1, 57–62.
- BOYT, W.P., MACKENZIE, P.K.I., PILSON, R.D. & LEAVIS, H., 1972. The importance of the donkey (Equus asinus) as a source of food and a reservoir of trypanosomes for Glossina morsitans Westw. Rhodesia Science News, 6, 18.
- BRUCE, D., 1895. Preliminary report on the tsetse fly disease or nagana in Zululand. Ubombo, Zululand, December 1895. Durban: Bennett and David.
- BUNGENER, W. & MULLER, G., 1976. Adharenzphanomene bei Trypanosome congolense. Zeitschrift für Tropenmedizin und Parasitologie, 27, 370–371.
- CHITAMBO, H. & ARAKAWA, A., 1991. Therapeutic effect of Berenil and Samorin in mice infected with four trypanosome populations isolated from Zambian cattle. Veterinary Parasitology, 39, 42–52.
- CHIZYUKA, H.G.B. & LUGURU, S.M.K., 1986. Dipping to control vectors of cattle parasites. Parasitology Today, 2, 123.
- CLARKSON, M.J. & MCCABE, W., 1970. Trypanosoma vivax in ruminants. Transactions of the Royal Society of Tropical Medicine and Hygiene, 64, 164–165.
- CLAUSEN, P.H., ADEYEMI, I., BAUER, B., BRELOEER, M., SALCHOW, F. & STAAK, C., 1998. Host preferences of tsetse (Diptera: Glossinidae) based on bloodmeal identifications. Medical and Veterinary Entomology, 12, 169–180.
- CLOUGH, O., 1929. Report on African Affairs for the Year 1929. Vol. I. London: Harrison and Sons Ltd.
- CLOUGH, O., 1931. Report on African Affairs for the Year 1931. Vol. III. Guildford and Esher: Billing and Sons Ltd.
- CLOUGH, O., 1932. Report on African Affairs for the Year 1932. Vol. IV. Guildford and Esher: Billing and Sons Ltd.
- CONNOR, R.J., 1989. Final Report of the Regional Trypanosomiasis Expert. Regional Tsetse and Trypanosomiasis Control Programme, Malawi, Mozambique, Zambia and Zimbabwe. December 1989. FGU-Kronberg Consulting and Engineering GmbH. Konigstein, West Germany.
- CONNOR, R.J. & HALLIWELL, R.W., 1987. Bovine trypanosomiasis in southern Tanzania: Parasitological and serological survey of prevalence. Tropical Animal Health and Production, 19, 165–172.
- CONNOR, R.J. & MUKANGI, D.J.A., 1986. Concurrent outbreak of pseudo-lumpy skin disease and acute Trypanosoma vivax infection in cattle. Tropical Animal Health and Production, 18, 127–132.
- CONNOR, R.J., MUKANGI, D.J.A. & HALLIWELL, R.W., 1989. Bovine trypanosomiasis in southern Tanzania: Investigation into the incidence of infection and duration of chemoprophylaxis. Tropical Animal Health and Production, 21, 135–140.
- CORTEN, J.J.F.M., TER HUURNE, A.A.H.M., MOORHOUSE, P.D.S. & DE ROOIJ, R.C., 1988. Prevalence of trypanosomiasis in cattle in South-west Zambia. Tropical Animal Health and Production, 20, 78–84.
- CROFT, S.L., EAST, J.S. & MOLYNEUX, D.H., 1982. Anti-trypanosomal factor in the haemolymph of Glossina. Acta Tropica, 39, 293–302.
- CUNNINGHAM, M.P., 1966. Immunity in bovine trypanosomiasis. East African Medical Journal, 43, 394–397.
- CUNNINGHAM, M.P. & VAN HOEVE, K., 1965. Diagnosis of trypanosomiasis in cattle. In: International Scientific Committee for Trypanosomiasis Research. Proceedings of the Tenth Meeting, Kampala 1964. Hertford, England: Stephen Austin & Sons Ltd.
- D’AMICO, F., POUSSINGA, J.M., LE MASSON, A. & CUISANCE, D.,1995. Pratiques pastorales Mboror et trypanosomoses bovines dans une zone de savanes humides de Centrafrique. Revue d’Elevage et de Médecine vétérinaire des Pays tropicaux, 48, 203–212.
- DALE, C., WELBURN, S.C., MAUDLIN, I. & MILLIGAN, P.J.M., 1995. The kinetics of maturation of trypanosome infections in tsetse. Parasitology, 111, 187–191.
- DARGIE, J.D., 1982. The influence of genetic factors on the resistance of ruminants to gastrointestinal nematode and trypanosome infections. In: OWEN, D.G., (ed.). Animal Models in Parasitology. London: Macmillan.
- DAVISON, G., 1990. Regional Tsetse and Trypanosomiasis Control Programme. Republic of Malawi National Tsetse and Trypanosomiasis Survey 1987–1989. Final Report. Gitec Consult GmbH, Dusseldorf, Federal Republic of Germany.
- DEHOUX, J.P., DIAW, M. & BULDGEN, A. 1996. Observation d’une flambée de trypanosomose equine à Trypanosoma vivax en zone urbaine au Sénégal. Tropicultura, 14, 35–26.
- DESQUESNES, M., 1997. Evaluation of a simple PCR technique for the diagnosis of Trypanosoma vivax infection in the serum of cattle in comparison to parasitological techniques and antigen-enzymelinked- immunosorbent assay. Acta Tropica, 65,139–148, 1997.
- DIARRA, B., DIALL, O., GEERTS, S., KAGERUKA, P., LEMMOUCHI, Y., SCHACHT, E., EISLER, M.C., & HOLMES, P., 1998. Field evaluation of the prophylactic effect of an isometamidium sustained-release device against trypanosomiasis in cattle. Antimicrobial Agents and Chemotherapy, 42, 1012–1014.
- DOLAN, R.B., OKECH, G., ALUSHULA, H., MUTUGI, M., STEVENSON, P., SAYER, P.D. & NJOGU, A.R., 1990. Homidium bromide as a chemoprophylactic for cattle trypanosomiasis in Kenya. Acta Tropica, 47, 137–144.
- DOLAN, R.B., SAYER, P.D., ALUSHULA, H. & HEATH, B.R., 1988. Pyrethroid impregnated ear tags in trypanosomiasis control. Tropical Animal Health and Production, 20, 267–268.
- DORAN, M., 1998. Socio-economic survey of tsetse-free and tsetse-infested areas in the Eastern Province, Zambia, 1997. Regional Tsetse and Trypanosomosis Control Programme (RTTCP). Harare, Zimbabwe.
- DORAN, M., 1998. Benefit: Cost analysis of border tsetse control operations in Zimbabwe. Regional Tsetse and Trypanosomosis Control Programme (RTTCP), Harare, Zimbabwe.
- DOWLER, M.E., SCHILLINGER, D. & CONNOR, R.J., 1989. Notes on the routine intravenous use of isometamidium in the control of bovine trypanosomiasis on the Kenya Coast. Tropical Animal Health and Production, 21, 4–10.
- DOYLE, J.J., 1977. Antigenic variation in salivarian trypanosomes. In: PINO, J., MILLER, L. & MCKELVEY, J., (eds). Blood-borne Parasitic Diseases. New York: Plenum Press.
- DU TOIT, R., 1954. Trypanosomiasis in Zululand and the control of tsetse by chemical means. Onderstepoort Journal of Veterinary Research, 26, 317–387.
- DWINGER, R.H., CLIFFORD, D.J., AGYEMANG, K., GETTINBY, G., GRIEVE, A.S., KORA, S., & BOJANG, M.A., 1993. Comparative studies on N’Dama and zebu cattle following repeated infections with Trypanosoma congolense. Research in Veterinary Science, 52, 292–298.
- DWINGER, R.H., AGYEMANG, K., SNOW, W.F., RAWLINGS, P., LEPERRE, P. & BAH, M.L., 1994. Productivity of trypanotolerant cattle kept under traditional management conditions in the Gambia. Veterinary Quarterly, 16, 81–86.
- DWINGER, R.H., GROOTENHUIS, J.G., MURRAY, M., MOLOO, S.K. & GETTINBY, G., 1986. Susceptibility of buffaloes, cattle and goats to infection with different stocks of Trypanosoma vivax transmitted by Glossina morsitans centralis. Research in Veterinary Science, 41, 307–315.
- DWINGER, R.H., LUCKINS, A.G., MURRAY, M., RAE, P. & MOLOO, S.K., 1986. Interference between different serodemes of Trypanosoma congolense in the establishment of superinfections in goats following cyclical transmission by tsetse. Parasite Immunology, 8, 293–305.
- DWINGER, R.H., LUCKINS, A.G., MURRAY, M., RAE, P. & MOLOO, S.K., 1989. Interference in the establishment of tsetse-transmitted Trypanosoma congolense, T. brucei or T. vivax superinfections in goats already infected with T. congolense or T. vivax. Veterinary Parasitology, 30, 177–189.
- DWINGER, R.H., MURRAY, M. & MOLOO, S.K., 1990. Parasite kinetics and cellular responses in goats infected and superinfected with Trypanosoma congolense transmitted by Glossina morsitans centralis. Acta Tropica, 47, 23–33.
- DWINGER, R.H., RUDIN, W., MOLOO, S.K. & MURRAY, M., 1988. Development of Trypanosoma congolense, T. vivax and T. brucei in the skin reaction induced in goats by infected Glossina morsitans centralis. Research in Veterinary Science, 44, 154–163.
- EISLER, M.C., AROWOLO, R.O.A., GAULT, E.A., MOLOO, S.K., HOLMES, P.H. & PEREGRINE, A.S., 1994. Isometamidium concentrations in the sera of Boran cattle: correlation with prophylaxis against tsetse-transmitted Trypanosoma congolense. Acta Tropica, 56, 39–50.
- EISLER, M.C., ELLIOTT, C.T., & HOLMES, P.H., 1996. A simple competitive enzyme immunoassay for the detection of the trypanocidal drug isometamidium. Therapeutic Drug Monitoring, 18, 73–79.
- EISLER, M.C., GAULT, E.A., SMITH, H.V., PEREGRINE, A.S. & HOLMES, P.H., 1993. Evaluation and improvement of an enzyme-linked immunosorbent assay for the detection of isometamidium in bovine serum. Therapeutic Drug Monitoring, 15, 236–242.
- ELCE, B.J., 1971. The transmission of Trypanosoma congolense through Glossina morsitans and the white mouse. Transactions of the Royal Society of Tropical Medicine and Hygiene, 65, 239.
- ELSEN, P., AMOUDI, M.A. & LECLERCQ, M., 1990. Première découverte de deux espèces de Glossines en Arabie Saoudite. Troisième Conference internationale des Entomologistes d’expression Française. Méthodes et responsabilités des Entomologistes d’aujourd’hui. Gembloux, 9–14 Juillet 1990.
- EMERY, D.L., BARRY, J.D. & MOLOO, S.K., 1980. The appearance of T. vivax in lymph following challenge of goats with infected Glossina morsitans morsitans. Acta Tropica, 37, 375–379.
- EMERY, D.L. & MOLOO, S.K., 1980. The sequential cellular changes in the local skin reaction produced in goats by Glossina morsitans morsitans infected with Trypanosoma (Trypanozoon) brucei. Acta Tropica, 37, 137–149.
- EPSTEIN, H., 1971. The Origin of the Domestic Animals of Africa. Vols. I & II. New York: Africana Publishing Corporation.
- EVANS, G., 1880. Report on ‘surra’ disease in the Dera Ismail Khan district. Punjab Government Military Department, No. 493–4467.
- FIENNES, R.N.T.-W., 1970. Pathogenesis and pathology of animal trypanosomiasis. In: mulligan, h.w. & potts, w.h., (eds). The African Trypanosomiases. London: George Allen and Unwin/Ministry of Overseas Development.
- FINELLE, P., 1973. Chimiothérapie et chimioprévention de trypanosomiase animale. Acquisitions récentes et situation actuelle. Les Cahiers de Medecine Veterinaire, 42, 215–226.
- FISON, T., 1985. Veterinary Investigation Centre, Mtwara, Tanzania. Unpublished data.
- FORD, J., 1971. The Role of the Trypanosomiases in African Ecology. Oxford: Clarendon Press.
- FRENCH, M.H., 1935. Nitrogen and mineral metabolism in Trypanosoma congolense disease. In: Annual Report of the Department of Veterinary Science and Animal Husbandry, Tanganyika, 1934.
- FRENCH, M.H. & HORNBY, H.E., 1936. Studies concerning the effect of the plane of nutrition on the course of animal trypanosomiasis. III. T. congolense disease from crisis to recovery. In: Annual Report of the Department of Veterinary Science and Animal Husbandry, Tanganyika, 1935.
- GARDINER, P.R., 1989. Recent studies of the biology of Trypanosoma vivax. Advances in Parasitology, 28, 229–317.
- GEERTS, S. & HOLMES, P.H., 1998. Drug management and parasite resistance in animal trypanosomiasis in Africa. PAAT technical and scientific series. Rome.
- GEERTS, S., KAGERUKA, P., DE DEKEN, R., BRANDT, J.R.A., KAZADI, J.M., DIARRA, B., EISLER, M.C., SCHACHT, E., & HOLMES, P.H., 1997. Prophylactic effects of isometamidium- and ethidium-sustained release devices against Trypanosoma congolense in cattle. Acta Tropica, 65, 23–31.
- GIBSON, W.C., 1985. Application of new technologies to epidemiology. British Medical Bulletin, 41, 115–121.
- GIBSON, W., 1994. Identification of trypanosomes in animals, humans and Glossina. Bulletin de la Societe de Pathologie Exotique, 87, 315–318.
- GINGRICH, J.B., MACKEN, L.M., JACKSON, P.R. & ROBERTS, D.R., 1985. Trypanosoma brucei: enhancement of infection rates in the tsetse fly, Glossina morsitans by feeding artificial bloodmeal mixtures. American Journal of Tropical Medical Hygiene, 34, 73–77.
- GOMBE, S., 1989. Endocrine effects of trypanosomiasis: Recent studies. Discovery and Innovation, 1, 30–33.
- GOMEZ, A.F., 1990. CTA/SADCC workshop on vector-borne diseases: Country report—Angola. A paper presented to the CTA/SADCC workshop on vector-borne diseases, Mbabane, Swaziland, 30 July 1990– 3 August 1990.
- GOODWIN, L.G., 1970. The pathology of African trypanosomiasis. Transactions of the Royal Society of Tropical Medicine and Hygiene, 64, 797–817.
- GRAY, A.R., 1974. Trypanosomal antigens and the control of trypanosomiasis. In: Control Programs for Trypanosomes and their Vectors. Actes du Colloque, Paris, 12–15 March 1974. Institut d’Élevage et de Médecine Vétérinaire des Pays Tropicaux.
- GRAY, A.R. & LUCKINS, A.G., 1976. Antigenic variation in salivarian trypanosomes. In: LUMSDEN, W.H.R. & EVANS, D.A., (eds). Biology of the Kinetoplastids. Vol. 1. London: Academic Press.
- GRAY, A.R. & ROBERTS, C.J., 1971. The cyclical transmission of strains of Trypanosoma congolense and T. vivax resistant to normal therapeutic doses of trypanocidal drugs. Parasitology, 63, 67–89.
- GRAY, A.R. & ROBERTS, C.J., 1971. The stability of resistance to diminazene aceturate and quinapyramine in a strain of Trypanosoma vivax during antelope. Parasitology, 63, 163–168.
- GREINER, M., KUMAR, S., & KYESWA, C., 1997. Evaluation and comparison of antibody ELISAs for serodiagnosis of bovine trypanosomosis. Veterinary Parasitology, 73, 197–205.
- GRIFFIN, L., 1978. African trypanosomiasis in sheep and goats. A review. The Veterinary Bulletin, 42, 819–825.
- GRIFFIN, L. & ALLONBY, E.W., 1979. Trypanotolerance in breeds of sheep and goats with an experimental infection of Trypanosoma congolense. Veterinary Parasitology, 5, 97–105.
- GRIFFIN, L., ALLONBY, E.W. & PRESTON, J.M., 1981. The interaction of Trypanosoma congolense and Haemonchus contortus infections in two breeds of goat. I. Parasitology. Journal of Comparative Pathology, 91, 85–95.
- GRIFFIN, L., WAGHELA, S. & ALLONBY, E.W., 1980. The immunosuppressive effects of experimental T. congolense infections in goats. Veterinary Parasitology, 7, 11–18.
- GROOTENHUIS, J.G., DWINGER, R.H., DOLAN, R.B., MOLOO, S.K. & MURRAY, M., 1990. Susceptibility of African buffalo and Boran cattle to Trypanosoma congolense transmitted by Glossina morsitans centralis. Veterinary Parasitology, 35, 219–231.
- HARLEY, J.M.B., 1966. Further studies on age and trypanosome infection rates in Glossina pallidipes Aust., G. palpalis fuscipes Newst. and G. brevipalpis Newst. in Uganda. Bulletin of Entomological Research, 57, 459–477.
- HENDERSON, W., 1931. The relationship of pica in cattle to trypanosomiasis. The Veterinary Journal, 87, 518–523.
- HENDY, C.R.C., 1988. The effects of trypanosomiasis prophylaxis and antihelmintic treatment in goats under traditional management in southern Tanzania. In: Livestock Production in Tsetse affected areas of Africa, Proceedings of a Meeting held 23–27 November 1987, Nairobi, Kenya ILCA/ILRAD. Nairobi: English Press.
- HENNING, M.W., 1956. Animal Diseases in South Africa. 3rd edn. Pretoria: Central News Agency Ltd.
- HOARE, C.A., 1972. The Trypanosomes of Mammals: A Zoological Monograph. Oxford and Edinburgh: Blackwell Scientific Publications.
- HOLMES, P.H., MAMMO, E., THOMSON, A., KNIGHT, P.A., LUCKEN, R., MURRAY, P.K., MURRAY, M., JENNINGS, F.W. & URQUHART, G.M., 1974. Immunosuppression in bovine trypanosomiasis. The Veterinary Record, 95, 86–87.
- HOLMES, P.H. & SCOTT, J.M., 1982. Chemotherapy against animal trypanosomiasis. In: BAKER, J.R., (ed.). Perspectives in Trypanosomiasis Research. Proceedings of the Twenty First Trypanosomiasis Seminar, London 24 September 1981, Letchworth: Research Studies Press.
- HOLMES, P.H., WHITELAW, D.D., BELL, I., MOLOO, S.K., HIRUMI, H., MURRAY, M. & URQUHART, G.M., 1985. The association between Samorin chemoprophylaxis and immune responses in cattle under experimental metacyclic Trypanosoma congolense challenge. The Proceedings of the International Scientific Council for Trypanosomiasis Research and Control, OAU/STRC, 18th Meeting, Harare, Zimbabwe 1985. Publication No. 113.
- HOPKINS, J.S., CHITAMBO, H., MACHILA, N., LUCKINS, A.G., RAE, P.F., VAN DEN BOSSCHE, P. & EISLER, M.C., 1998. Adaptation and validation of the antibody trapping ELISA using dried blood spots on filter paper, for epidemiological surveys of tsetse transmitted trypanosomosis in cattle. Preventive Veterinary Medicine, 37, 91–99.
- HORNBY, H.E., 1921. Trypanosomes and trypanosomiasis of cattle. Journal of Comparative Pathology, 34, 211–240.
- HORNBY, H.E., 1952. Animal Trypanosomiasis in East Africa, 1949. London, HMSO.
- HORNBY, H.E. & BAILEY, H.W., 1932. Research on trypanosomiasis. In: Annual Report of the Department of Veterinary Science and Animal Husbandry, Tanganyika, 1931Annual Report of the Department of Veterinary Science and Animal Husbandry, Tanganyika, 1931..
- IBRAHIM, E.A.R., INGRAM, G.A. & MOLYNEUX, D.H., 1984. Haemagglutinins and parasite agglutinins in haemolymph and gut of Glossina. Tropical Medicine and Parasitology, 35, 151–156.
- IKEDE, B.O., 1979. Genital lesions in experimental chronic Trypanosoma brucei infection in rams. Research in Veterinary Science, 26, 145–151.
- IKEDE, B.O., ELHASSAN, E. & AKPAVIE, S.O., 1988. Reproductive disorders in African trypanosomiasis: A review. Acta Tropica, 45, 5–10.
- IKEDE, B.O. & LOSOS, G.J., 1972. Pathology of the disease in sheep produced experimentally by Trypanosoma brucei. Veterinary Pathology, 9, 278– 289.
- ILCA, 1979. Small Ruminant Production in the Humid Tropics. ILCA Systems Study 3. Addis Ababa: International Livestock Centre for Africa.
- ILRAD, 1983. ilrad, 1982. Annual Report of the International Laboratory for Research on Animal Diseases. Nairobi: ILRAD.
- ILRAD, 1989. ilrad1988: Annual Report of the International Laboratory for Research on Animal Diseases. Nairobi, ILRAD.
- IMBUGA, M.O., OSIR, E.O. & LABONGO, V.L., 1992. Inhibitory effect of Trypanosoma brucei brucei on Glossina morsitans midgut trypsin in vitro. Parasitology Research, 78, 273–276.
- IMBUGA, M.O., OSIR, E.O., LABONGO, V.L., DARJI, N. & OTIENO, L.H., 1992. Studies on tsetse midgut factors that induce differentiation of bloodstream Trypanosoma brucei brucei in vitro. Parasitology Research, 78, 10–15.
- JACKSON, P.R. & DIGGS, C.L., 1983. Trypanosoma rhodesiense bloodstream trypomastigotes and culture procyclic cell surface carbohydrates. Journal of Protozoology, 30, 662–668.
- JACKSON, P.R., HONIGBERG, B.M. & HOLT, S.C., 1978. Lectin analysis of Trypanosoma congolense bloodstream trypomastigote and culture procyclic surface saccarides by agglutination and electron microscopic techniques. Journal of Protozoology, 25, 471–481.
- JEFFRIES, D., HELFRICH, M.P. & MOLYNEUX, D.H., 1987. Cibarial infections of Trypanosoma vivax and T. congolense in Glossina. Parasitology Research, 73, 289–292.
- JEMAL, A. & HUGH-JONES, M.E., 1995. Association of tsetse control with health and productivity of cattle in the Didessa Valley, western Ethiopia. Preventive Veterinary Medicine 22, 29–40.
- JENNI, L., MARTI, S., SCHWEIZER, J., BETSCHART, E., LE PAGE, R.W.F., WELLS, J.M., TAIT, A., PAINDAVOINE, P., PAYS, E. & STEINERT, M., 1986. Hybrid formation between African trypanosomes during cyclical transmission. Nature, 322, 173–175.
- JENNI, L., MOLYNEUX, D.H., LIVESEY, J.L. & GALUN, R., 1980. Feeding behaviour of tsetse flies infected with salivarian trypanosomes. Nature, 283, 383–385.
- JONES-DAVIES, W.J., 1968. Diminazine aceturate and homidium chloride resistance in tsetse fly-transmitted trypanosomes of cattle in northern Nigeria. The Veterinary Record, 83, 433–437.
- JORDAN, A.M., 1976. Tsetse flies as vectors of trypanosomes. Veterinary Parasitology, 2, 143–152.
- JORDAN, A.M., 1986. Trypanosomiasis Control and African Rural Development. London and New York: Longman.
- JOSHUA, R.A. & KAYIT, Y.S., 1984. The course of fatal Trypanosoma simiae infection in domestic sheep. Acta Tropica, 41, 91–92.
- JOSHUA, R.A., OBWOLO, M.J., BWANGAMOI, O. & MANDEBVU, E., 1995.Resistance to diminazene aceturate by Trypanosoma congolense from cattle in the Zambezi Valley of Zimbabwe. Veterinary Parasitology, 60, 1–6.
- KAAYA, G.P. & ODUOR-OKELO, D., 1980. The effects of Trypanosoma congolense infection on the testis and epididymis of the goat. Bulletin of Animal Health and Production in Africa, 28, 1–5.
- KAMINSKY, R., 1990. In vitro techniques for assessment of drug resistance in trypanosomes. AgBiotech News and Information, 2, 205–210.
- KANYARI, P.W.N., ALLONBY, E.W., WILSON, A.J. & MUNYUA, W.K., 1983. Some economic effects of trypanosomiasis in goats. Tropical Animal Health and Production, 15, 153–160.
- KAPPMEIER, K., NEVILL, E.M. & BAGNALL, R.J., 1998. Review of tsetse and trypanosomosis in South Africa. Onderstepoort Journal of Veterinary Research, 65, 195-203.
- KATAKURA, K., LUBINGA, C., CHITAMBO, H. & TRADA, Y., 1997. Detection of Trypanosoma congolense and T. brucei subspecies in cattle in Zambia by polymerase chain reaction from blood collected on a filter paper. Parasitology Research, 83, 241–245.
- KATUNGUKA-RWAKISHAYA, E., 1996. The prevalence of trypanosomosis in small ruminants and pigs in a sleeping sickness endemic area of Buikwe County, Mukono district, Uganda. Revue d’Elevage et de Médecine Vétérinaire des Pays tropicaux, 49, 56–58.
- KATUNGUKA-RWAKISHAYA, E., PARKINS, J.J., FISHWICK, G., MURRAY, M., & HOLMES, P.H., 1995. The influence of energy intake on the pathophysiology of Trypanosoma congolense infection in Scottish Blackface sheep. Veterinary Parasitology, 59, 207–218.
- KATENDE, J.M., MUSOKE, A.J., NANTULYA, V.M. & GODDEERIS, B.M., 1987. A new method for fixation and preservation of trypanosomal antigens for use in the indirect immunoflourescence antibody test for diagnosis of bovine trypanosomosis. Tropical Medicine and Parasitology, 38, 41–44.
- KELLEY, S. & SCHILLINGER, D., 1983. Improved field diagnostic technique for trypanosomiasis by use of a minicentrifuge. The Veterinary Record, 113, 219.
- KILLICK-KENDRICK, R., 1968. The diagnosis of trypanosomiasis in livestock: A review of current techniques. The Veterinary Bulletin, 38, 191–197.
- KINABO, L.D.B., 1993. Pharmacology of existing drugs for animal trypanosomiasis. Acta Tropica, 54, 169–183.
- KINABO, D.B. & BOGAN, J.A., 1988. The pharmacology of isometamidium. Journal of Veterinary Pharmacology and Therapeutics, 11, 233–245.
- KNOWLES, G., ABEBE, G. & BLACK, S.J., 1989. Detection of parasite peptidase in the plasma of heifers infected with Trypanosoma congolense. Molecular and Biochemical Parasitology, 34, 25–34.
- KOMOIN-OKA, C., TRUC, P., BENGALY, Z., FORMENTY, P., DUVALLET, G., LAUGINIE, F., RAATH, J.P., N’DEPO, A.E. & LEFORBAN, Y., 1994. Etude de la prévalence des infections à trypanosomes chez différent espèces d’animaux sauvages du parc national de la Comoé en Côte d’Ivoire: résultats préliminaires sur la comparison de trois méthodes de diagnostic. Revue d’Elevage et de Médecine Vétérinaire des Pays tropicaux, 47,189–194.
- KRATZER, R.D. & ONDIEK, F.O., 1989. The buffy coat double centrifugation technique, an improved method for the diagnosis of African trypanosomosis. The Proceedings of the International Scientific Council for Trypanosomosis Research and Control, OAU/ISCTRC, 20th Meeting, Mombassa, Kenya 1989. Publication No. 115.
- LANHAM, S.M. & GODFREY, D.G., 1970. The isolation of salivarian trypanosomes from man and other mammals using DEAE-cellulose. Experimental Parasitology, 28, 521–534.
- LAVERAN, A. & MESNIL, F., 1904. Trypanosomes and Trypanosomiases. English translation and enlargement, Nabarro, D., 1907. London: BalliTre, Tindall & Co.
- LAWRENCE, J.A., FOGGIN, C.M. & NORVAL, R.A.I., 1980. The effect of war on the control of diseases of livestock in Rhodesia (Zimbabwe). The Veterinary Record, 107, 82–85.
- LEACH, T.M. & ROBERTS, C.J., 1981. Present status of chemotherapy and chemoprophylaxis of animal trypanosomiasis in the eastern hemisphere. Pharmacology and Therapeutics, 13, 91–147.
- LEAK, S.G.A. & ROWLANDS, G.J., 1997. The dynamics of trypanosome infections in natural population of tsetse (Diptera: Glossinidae) studied using wing-fray and ovarian ageing techniques. Bulletin of Entomological Research, 87, 273–282.
- LEEFLANG, P., 1975. The predominance of Trypanosoma vivax infections of cattle at a distance from savannah tsetse concentration. Tropical Animal Health and Production, 7, 201–204.
- LEWIS, A.R. & THOMSON, J.W., 1974. Observations on an isometamidium resistant strain of Trypanosoma congolense in Rhodesia. Rhodesian Veterinary Journal, 4, 62–67.
- LLEWELYN, C.A., LUCKINS, A.G., MUNRO, C.D. & PERRIE, J., 1987. The effect of Trypanosoma congolense infection on the oestrous cycle of the goat. British Veterinary Journal, 143, 423–431.
- LLEWELYN, C.A., MUNRO, C.D., LUCKINS, A.G., JORDT, T., MURRAY, M. & LORENZINI, E., 1988. The effects of Trypanosoma congolense infection on the oestrous cycle of the Boran cow. British Veterinary Journal, 144, 379–387.
- LOGAN, L.L., GOODWIN, J.T., TEMBELY, S. & CRAIG, T.M., 1984. Maintaining Zebu Maure cattle in a tsetse infested area of Mali. Tropical Animal Health and Production, 16, 1–12.
- LOSOS, G.J., 1986. Infectious Tropical Diseases of Domestic Animals. Longman Scientific and Technical in association with the International Development and Research Centre, Canada.
- LOSOS, G. & CHOUINARD, A., 1979. Pathogenicity of trypanosomes. Proceedings of a Workshop held at Nairobi, Kenya, 20–23 November 1978. Ottawa: International Development Research Centre.
- LOSOS, G.J., PARIS, J., WILSON, A.J. & DAR, F.K., 1973. Pathology of the disease in cattle caused by Trypanosoma congolense. Bulletin of Epizootic Diseases of Africa, 21, 239–248.
- LUCKINS, A.G., 1977. Detection of antibodies in trypanosome-infected cattle by means of a microplate enzyme-linked immunosorbent assay. Tropical Animal Health and Production, 9, 53–62.
- LUCKINS, A.G., 1979. Trypanosomes in the lymph nodes of cattle and sheep infected with Trypanosoma congolense. Research in Veterinary Science, 27, 129–131.
- LUCKINS, A.G. & MEHLITZ, D., 1978. Evaluation of an indirect fluorescent antibody test, enzyme-linked immunosorbent assay and quantification of immunoglobulins in the diagnosis of bovine trypanosomiasis. Tropical Animal Health and Production, 10, 149–159.
- LUCKINS, A.G., SUTHERLAND, D., MWANGI, D. & HOPKINS, J., 1994. Early stages of infection with Trypanosoma congolense: parasite kinetics and expression of metacyclic variable antigen types. Acta Tropica, 58, 199–206.
- LUMSDEN, W.H.R., KIMBER, C.D., EVANS, D.A. & DOIG, S.J., 1979. Trypanosoma brucei: Miniature anion-exchange centrifugation technique for detection of low parasitaemias: Adaptation for field use. Transactions of the Royal Society of Tropical Medicine and Hygiene, 73, 312–317.
- MACKENZIE, P.K.I., BOYT, W.P. & EMSLIE, V.W., 1975. Two cases of natural infection of sheep with Trypanosoma (Nannomonas) simiae (Bruce and others 1912). Rhodesian Veterinary Journal, 6, 8–14.
- MACKENZIE, P.K.I., BOYT, W.P., EMSLIE, V.W., LANDER, K.P. & SWANEPOEL, R., 1975. Immunosuppression in ovine trypanosomiasis. The Veterinary Record, 97, 452–453.
- MACLENNAN, K.J.R., 1970. The epizootiology of trypanosomiasis in livestock in West Africa. In: mulligan, h.w. & potts, w.h., (eds). The African Trypanosomiases. London: George Allen and Unwin/Ministry of Overseas Development.
- MACLENNAN, K.J.R., 1974. The epizootiology of tsetse transmitted trypanosomiasis in relation to livestock development and control measures. In: Control Programmes for Trypanosomes and their Vectors. Actes du Colloque, Paris 12–15 March 1974, Institut d’Élevage et de Médecine Vétérinaire des Pay Tropicaux.
- MACLENNAN, K.J.R., 1975. A review of operations against tsetse flies and trypanosomiasis in Zambia and recommendations for the future. Report to the Director of Veterinary and Tsetse Control Services, Lusaka, Zambia.
- MACLENNAN, K.J.R., 1980. Tsetse-transmitted trypanosomiasis in relation to the rural economy in Africa. I. Tsetse infestation. World Animal Review, 36, 2–17.
- MACLENNAN, K.J.R. & NA’ISA, B.K., 1970. Relapsing Trypanosoma vivax infections in Nigerian zebu cattle treated with diminazene aceturate. Tropical Animal Health and Production, 2, 189–195.
- MAGONA, J.W., KATABAZI, W., OLAHO-MUKANI, W., MAYENDE, J.S.P. & WALUBENGO, J., 1997. Haemorrhagic Trypanosoma vivax outbreak in cattle in Mbale and Tororo districts in eastern Uganda. Journal of Protozoology Research, 7, 48–53.
- MAKUMYAVIRI, A.M., 1990. The epidemiological value of experimental host and animal reservoir of Trypanosoma brucei gambiense. Revue de Médecine Vétérinaire, 141, 873–875.
- MAKUMYAVIRI, A., MEHLITZ, D., KAGERUKA, P., KAZYUMBA, G.L. & MOLISHO, D., 1989. Le réservoir animal de Trypanosoma brucei gambiense au Zaire: Infections trypanosomiennes dans deux foyer du Bas-Zaire. Tropical Medicine and Parasitology, 40, 258–262.
- MALOO, S.H., CHEMA, S., CONNOR, R., DURKIN, J., KIMOTHO, P., MAEHL, J.H.H., MUKENDI, F., MURRAY, M., RARIEYA, J.M. & TRAIL, J.C.M., 1988. The use of chemoprophylaxis in East African Zebu village cattle exposed to trypanosomiasis in Muhaka, Kenya. In: Livestock Production in Tsetse affected areas of Africa. Proceedings of a Meeting of the African Trypanotolerant Livestock Network, 23–27 November 1987, Nairobi, Kenya. Nairobi: English Press.
- MASAKE, R.A., 1980. The pathogenesis of infection with Trypanosoma vivax in goats and cattle. The Veterinary Record, 107, 551–557.
- MASAKE, R.A., NANTULYA, V.M., AKOL, G.W.O. & MUSOKE, A.J., 1984. Cerebral trypanosomiasis in cattle with mixed Trypanosoma congolense and T. brucei brucei infections. Acta Tropica, 41, 237–246.
- MAUDLIN, I., KABAYO, J.P., FLOOD, M.E.T. & EVANS, D.A., 1984. Serum factors and the maturation of Trypanosoma congolense infections in Glossina morsitans. Zeitschrift für Parasitenkunde, 70, 11–19.
- MAUDLIN, I. & WELBURN, S.C., 1987. Lectin mediated establishment of midgut infections of Trypanosoma congolense and Trypanosoma brucei in Glossina morsitans. Tropical Medicine and Parasitology, 38, 167–170.
- MAUDLIN, I. & WELBURN, S.C., 1988. The role of lectins and trypanosome genotype in the maturation of midgut infections in Glossina morsitans. Tropical Medicine and Parasitology, 39, 56–58.
- MATSON, B.A., 1959. An investigation into the livestock problems of the Lower Shire Valley of Nyasaland with particular reference to the disease trypanosomiasis. Department of Veterinary Services and Animal Industry, Nyasaland.
- MCCULLY, R.M. & NEITZ, W.O., 1971. Clinicopathological study on experimental Trypanosoma brucei infections in horses. Part 2. Histopathological findings in the nervous system and other organs of treated and untreated horses reacting to nagana. Onderstepoort Journal of Veterinary Research, 38, 141–176.
- MIHOK, S., OLUBAYO, R.O., DARJI, N. & ZWEYGARTH, E., 1993. The influence of host blood on infection rates in Glossina morsitans spp. infected with Trypanosoma congolense, T. brucei and T. simiae. Parasitology, 107, 41–48.
- MILLIGAN, P.J.M. & BAKER, R.D., 1988. A model of tsetse-transmitted animal trypanosomiasis. Parasitology, 96, 211–239.
- MIMBA, D. & EVANS, H.G., 1990. Zamanglo Industrial Corporation Limited, Lusaka, Zambia. Unpublished data.
- MOLOO, S.K., 1993. The distribution of Glossina species in Africa and their natural hosts. Insect Science and its Application, 14, 511–527.
- MOLOO, S.K., 1981. Effects of maintaining Glossina morsitans morsitans on different hosts upon the vector’s subsequent infection rates with pathogenic trypanosomes. Acta Tropica, 38, 125–136.
- MOLOO, S.K., KUTUZA, S.B. & BOREHAM, P.F.L., 1980. Studies on Glossina pallidipes, G. fuscipes fuscipes and G. brevipalpis in terms of the epidemiology and epizootiology of trypanosomiases in South Eastern Uganda. Annals of Tropical Medicine and Parasitology, 74, 219–237.
- MOLYNEUX, D.H. & ASHFORD, R.W., 1983. The Biology of Trypanosoma and Leishmania Parasites of Man and Domestic Animals. London: Taylor and Francis.
- MOLYNEUX, D.H. & JEFFERIES, D., 1986. Feeding behaviour of pathogen-infected vectors. Parasitology, 92, 721–726.
- MORRISON, W.I., MURRAY, M. & AKOL, G.W.O., 1985. Immune responses of cattle to African trypanosomes. In: TIZARD, I., (ed.). Immunology and Pathogenesis of Trypanosomiasis. Boca Raton, Florida: CRC Press Ltd.
- MORRISON, W.I., MURRAY, M. & SAYER, P.D., 1979. Pathogenesis of tissue lesions in T. brucei infections. In: LOSOS, G. & CHONINARD, A., (eds). Pathogenicity of Trypanosomes. Proceedings of a Workshop held at Nairobi, Kenya, 20–23 November 1978. Ottawa: International Development Research Centre.
- MORRISON, W.I., WELLS, P.W., MOLOO, S.K., PARIS, J. & MURRAY, M., 1982. Interference in the establishment of superinfections with Trypanosoma congolense in cattle. Journal of Parasitology, 68, 755–764.
- MOSER, D.R., COOK, G.A., OCHS, D.E. & BAILEY, C.P., 1989. Detection of T. congolense and T. brucei subspecies by DNA amplification using the polymerase chain reaction. Parasitology, 99, 57–66.
- MULLA, A.F. & RICKMAN, L.R., 1988. How do African game animals control trypanosome infections? Parasitology Today, 4, 352–354.
- MULLA, A.F. & RICKMAN, L.R., 1988. Evidence for the presence of an innate trypanosomicidal factor in the serum of a non-immune African Waterbuck (Kobus ellipsiprymnus). Transactions of the Royal Society of Tropical Medicine and Hygiene, 82, 97–98.
- MULLIGAN, H.W. & POTTS, W.H., 1970. The African Trypanosomiasis. London: George Allen and Unwin/Ministry of Overseas Development.
- MURRAY, M., 1974. The pathology of African trypanosomiases. In: BRENT, L. & HOLBORROW, T., (eds). Progress in Immunology. II. Vol. 4. Amsterdam: North-Holland Publishing Company.
- MURRAY, M., BARRY, J.D., MORRISON, W.I., WILLIAMS, R.O., HIRUMI, H. & ROVIS, L., 1979. A review of the prospects for vaccination in African trypanosomiasis. Part I. World Animal Review, 32, 9–13.
- MURRAY, M. & BLACK, S.J., 1985. African trypanosomiasis in cattle: Working with nature’s solution. Veterinary Parasitology, 18, 167–182.
- MURRAY, M., CLIFFORD, D.J., GETTINBY, G., SNOW, W.F. & MCINTYRE, W.I.M., 1981. Susceptibility to African trypanosomiasis of N’Dama and Zebu cattle in an area of Glossina morsitans submorsitans challenge. The Veterinary Record, 109, 503–510.
- MURRAY, M. & DEXTER, T.M., 1988. Anaemia in bovine African trypanosomiasis. Acta Tropica, 45, 389–432.
- MURRAY, M., MORRISON, W.I., MURRAY, P.K., CLIFFORD, D.J. & TRAIL, J.C.M., 1979. Trypanotolerance—a review. World Animal Review, 31, 2–12.
- MURRAY, M., MORRISON, W.I. & WHITELAW, D.D., 1982. Host susceptibility to African trypanosomiasis: Trypanotolerance. Advances in Parasitology, 21, 1–68.
- MURRAY, M., MURRAY, P.K. & MCINTYRE, W.I.M., 1977. An improved parasitological technique for the diagnosis of African trypanosomiasis. Transactions of the Royal Society of Tropical Medicine and Hygiene, 71, 325–326.
- MURRAY, M., TRAIL, J.C.M. & D’IETEREN, G.D.M., 1990. Trypanotolerance in cattle and prospects for the control of trypanosomiasis by selective breeding. Revue Scientifique et Technique de L’Office International des Épizooties, 9, 369–386.
- MURILLA, G.A., MDACHI, R.E. & KARANJA, W.M., 1996. Pharmacokinetics, bioavailability and tissue residues of [14C]isometamidium in non-infected and Trypanosoma congolense-infected Boran cattle. Acta Tropica, 61, 277–292.
- MUTAYOBA, B.M. & GOMBE, S., 1989. Effects of African trypanosomiasis on plasma cortisol and thyroxine concentration in goats. Research in Veterinary Science, 47, 315–318.
- MUTAYOBA, B.M., GOMBE, S., KAAYA, G.P. & WAINDI, E.N., 1988. Trypanosome-induced ovarian dysfunction. Evidence of higher residual fertility in trypanotolerant small East African goats. Acta Tropica, 45, 225–237.
- MUTAYOBA, B.M., GOMBE, S., WAINDI, E.N. & KAAYA, G.P., 1989. Comparative trypanotolerance of the small East African breed of goats from different localities to Trypanosoma congolense infection. Veterinary Parasitology, 31, 96–105.
- MUTAYOBA, B.M., MEYER, H.H.D., OSASO, J. & GOMBE, S., 1989. Trypanosome-induced increase in prostaglandin-F-2-alpha and its relationship with corpus luteum function in the goat. Theriogenology, 32, 545–556.
- MUTAYOBA, B.M., O’HARAOIRERI, H.B. & GOMBE, S., 1988. Trypanosomeinduced depression of plasma thyroxine levels in prepubertal and adult female goats. Acta Endocrinologica (Copenhagen), 119, 21–26.
- MUTHARIA, L.M. & PEARSON, T.W., 1987. Surface carbohydrates of procyclic forms of African trypanosomes studied using fluorescence activated cell sorter analysis and agglutination of lectins. Molecular and Biochemical Parasitology, 23, 165–172.
- MWAMBU, P.M. & LOSOS, G.J., 1979. Ultrastructural changes in blood vessels of tissues of cattle experimentally infected with Trypanosoma congolense and T. vivax: A preliminary report. In: LOSOS, G. & CHOUINARD, A., (eds). Pathogenicity of Trypanosomes. Proceedings of a Workshop held at Nairobi, Kenya, 20–23 November 1978. Ottawa: International Development Research Centre.
- NANTULYA, V.M., 1986. Immunological approaches to the control of animal trypanosomiasis. Parasitology Today, 2, 168–173.
- NANTULYA, V.M., 1990. Trypanosomiasis in domestic animals: the problems of diagnosis. Revue Scientifique et Technique de L’Office International des Épizooties, 9, 357–367.
- NANTYLYA, V.M., DOYLE, J.J. & JENNI, L., 1978. Studies on Trypanosoma (Nannomonas) congolense. I. On the morphological appearance of the parasite in the mouse. Acta Tropica, 35, 329–337.
- NANTYLYA, V.M. & LINDQUIST, K.J., 1989. Antigen-detection enzyme immunoassays for the diagnosis of Trypanosoma vivax, T. congolense and T. brucei infections in cattle. Tropical Medicine and Parasitology, 40, 267–272.
- NANTYLYA, V.M., LINDQUIST, K.J. & STEVENSON, P., 1989. Towards improved diagnosis of trypanosomiasis. In: Livestock Production and Diseases in the Tropics. Proceedings of the Sixth International Conference on Livestock Production and Disease in the Tropics held in Wegeningen, The Netherlands, August 28 – September 1, 1989. Vol. 2, In Press.
- NANTYLYA, V.M., MUSOKE, A.J., RURANGIRWA, F.R., SAIGAR, N. & MINJA, S.H., 1987. Monoclonal antibodies that distinguish Trypanosoma congolense, T. vivax and T. brucei. Parasite Immunology, 9, 421–431.
- NDAO, M., PANDEY, V.S., ZINSSTAG, J., PFISTER, K. & VAN MEIRVENNE, N., 1995. Evaluation of sodium dodecyl sulfate (SDS) as a haemolytic agent for the detection of microfilaria and trypanosomes in the blood of cattle. Annales de la Société Belge de Médicine Tropicale, 75,145–148.
- NDOUTAMIA, G., MOLOO, S.K., MURPHY, N.B. & PEREGRINE, A.S., 1993. Derivation and characterization of a quinapyramine-resistant clone of Trypanosoma congolense. Antimicrobial Agents and Chemotherapy, 37, 1163–1166.
- NGUU, E.K., OSIR, E.O., IMBUGA, M.O. & OLEMBO, N.K., 1996. The effect of host blood on the in vitro transformation of bloodstream trypanosomes by tsetse midgut homogenates. Medical and Veterinary Entomology, 10, 317–322.
- NJOGU, A.R., OLAHO, W.M., SAYER, P.D., ALUSHULA, H. & DOLAN, R.B., 1982. Strategic use of chemoprophylaxis against trypanosomiasis in different types of Boran cattle. Kenya Veterinarian, 6, 7.
- OGUNYEMI, O. & ILEMOBADE, A.A., 1989. Prophylaxis of African animal trypanosomiases. A review of some factors that may influence the duration of isometamidium chloride prophylaxis. Veterinary Bulletin, 59, 1–4.
- OGWU, D. & NJOKU, C.O., 1987. Effect of pregnancy on clinical manifestation of bovine trypanosomiasis. Veterinary Parasitology, 24, 25–33.
- OGWU, D., NJOKU, C.O. & OSORI, D.I.K., 1986. Effects of experimental Trypanosoma vivax infection on first-, second-, and third-trimester pregnancy in heifers. Theriogenology, 25, 383–398.
- OGWU, D. & NURU, S., 1981. Transplacental transmission of trypanosome in animals and man. A review. The Veterinary Bulletin, 51, 381–384.
- OKECH, G., WATSON, E.D., LUCKINS, A.G. & MAKAWITI, D.W., 1996. The effect of experimental infection of Boran cattle in early and mid-pregnancy with Trypanosoma vivax. British Veterinary Journa, 152, 441–451.
- OMEKE, B.C.O., 1994. Pig trypanosomosis: prevalence and significance in the endemic Middle Belt zone of Southern Nigeria. Revue d’Elevage et de Médecine Vétérinaire des Pays tropicaux 47, 381–386.
- PARIS, J., MURRAY, M. & MCODIMBA, F., 1982. A comparative evaluation of the parasitological techniques currently available for the diagnosis of African trypanosomiasis in cattle. Acta Tropica, 39, 307–316.
- PEREGRINE, A.S., 1994. Chemotherapy and delivery systems: haemoparasites. Veterinary Parasitology, 54, 223–248.
- PEREGRINE, A.S. & MAMMAN, M., 1993. Pharmacology of diminazene: a review. Acta Tropica, 54: 185–203.
- PEREGRINE, A.S., MOLOO, S.K. & WHITELAW, D.D., 1991. Differences in sensitivity of Kenyan Trypanosoma vivax populations to the prophylactic and therapeutic actions of isometamidium chloride in Boran cattle. Tropical Animal Health and Production, 23, 29–38.
- PEREGRINE, A.S., OGUNYEMI, O., WHITELAW, D.D., HOLMES, P.H., MOLOO, S.K., HIRUMI, H., URQUHART, G.M. & MURRAY, M., 1988. Factors influencing the duration of isometamidium chloride (Samorin) prophylaxis against experimental challenge with metacyclic forms of Trypanosoma congolense. Veterinary Parasitology, 28, 53–64.
- PILSON, R.D., BOYT, W.P. & MACKENZIE, P.K.I., 1978. The relative attractiveness of cattle, sheep and goats to Glossina morsitans morsitans Westwood and G. pallidipes (Diptera: Glossinidae) in the Zambezi valley of Rhodesia. Bulletin of Entomological Research, 68, 489–495.
- PINDER, M., BAUER, J., VAN MELICK, A. & FUMOUX, F., 1988. Immune responses of trypanoresistant and trypanosusceptible cattle after cyclic infection with Trypanosoma congolense. Veterinary Immunology and Immunopathology, 18, 245–257.
- REIFENBERG, J.M., SOLANO, P., BAUER, B., KABORE, I., CUNY, G., DUVALLET, G. & CUISANCE, D., 1997. Apport de la technique PCR pour une meilleure compréhension de l’épizootiologie des trypanosomoses bovines: exemple de la zone d’aménagement pastoral de Yalé au Burkina Faso. Revue d’Elevage et de Médecine vétérinaire des Pays tropicaux, 50, 14–22.
- RICKMAN, L.R. & KOLALA, F., 1982. Effects of some African game animal sera on Trypanosoma brucei rhodesiense and T. b. brucei clones. Tropenmedizin und Parasitologie, 33, 129–135.
- ROBERTS, L.W., 1981. Probing by Glossina morsitans and transmission of Trypanosoma congolense. American Journal of Tropical Medicine and Hygiene, 30, 948–951.
- ROBERTS, C.J., GRAY, M.A. & GRAY, A.R., 1969. Local skin reactions in cattle at the site of infection with Trypanosoma congolense by Glossina morsitans and G. tachinoides. Transactions of the Royal Society of Tropical Medicine and Hygiene, 63, 620–624.
- ROBERTSON, A.G., 1983. The feeding habits of tsetse flies in Zimbabwe (formerly Rhodesia) and their relevance to some tsetse control measures. Smithersia No. 1, 1–72.
- RODITI, I. & PEARSON, T.W., 1990. The procyclin coat of African trypanosomes (or the not-so-naked trypanosome). Parasitology Today, 6, 79–82.
- ROEDER, P., SCOTT, J.M. & PEGRAM, R.G., 1984. Acute Trypanosome vivax infection of Ethiopian cattle in the apparent absence of tsetse. Tropical Animal Health and Production, 16, 141–147.
- ROELANTS, G.E., 1986. Natural resistance to African trypanosomiasis. Parasite Immunology, 8, 1–10.
- ROELANTS, G.E., TAMBOURA, I., SIDIKI, D.B., BASSINGA, A. & PINDER, M., 1983. Trypanotolerance: An individual not a breed character. Acta Tropica, 40, 99–104.
- ROGERS, D.J., 1980. Epizootiology: The tsetse cattle interface. In: FAO Report of the Expert Consultation on Research on Trypanosomiasis, Rome, 1–5 October 1979. FAO AGA:Tryp/7G/25. Rome: Food and Agriculture Organization.
- ROGERS, D.J., 1985. Trypanosomiasis ‘risk’ or ‘challenge’: A review. Acta Tropica, 42, 5–23.
- ROGERS, D.J., 1988. A general model for the African trypanosomiasis. Parasitology, 97, 193–212.
- ROGERS, D.J. & BOREHAM, P.F.L., 1973. Sleeping sickness survey in the Serengeti area (Tanzania) 1971 II. The vector role of Glossina swynnertoni Austen. Acta Tropica, 30, 24–35.
- ROTTCHER, D. & SCHILLINGER, D., 1985. Multiple drug resistance in Trypanosoma vivax in the Tana River district of Kenya. The Veterinary Record, 177, 557–558.
- ROWLANDS, G.J., MULATU, W.A., AUTHIE, E., D’IETEREN, G.D.M., LEAK, S.G.A., PEREGRINE, A.S. & TRAIL, J.C.M., 1990. Prevalence of Trypanosoma congolense in East African Zebu cattle under high tsetse challenge. Proceedings of the Society of Veterinary Epidemiology and Preventive Medicine, Belfast 1990.
- RYAN, L., CROFT, S.L., EAST, J.S., MOLYNEUX, D.H. & BALDRY, D.A.T., 1982. Differences in rates of acquisition of trypanosome infections between Glossina species in the field. Annales de la Société Belge de Médicine Tropicale, 62, 291–300.
- SCOONES, I., 1995. Exploiting heterogeneity: habits used by cattle in dryland Zimbabwe. Journal of Arid Environments, 29, 221–237.
- SEEBECK, T., HEMPHILL, A. & LAWSON, D., 1990. The cytoskeleton of trypanosomes. Parasitology Today, 6, 49–52.
- SEED, J.R. & SECHELSKI, J.B., 1989. Mechanisms of long slender (LS) to short stumpy (SS) transformation in the African trypanosomes. The Journal of Protozoology, 36, 572–576.
- SEHOF, C.F.H., 1975. Tsetse bloodmeal collections from the Kakumbi area of the Luangwa Valley (1973–1974).
- SEKONI, V.O., KUMI-DIAKA, J., SAROR, D. & NJOKU, C., 1988. The effect of Trypanosoma vivax and Trypanosoma congolense infections on the reaction time and semen characteristics in the Zebu bull. British Veterinary Journal, 144, 388–394.
- SHARPE, R.T., LANGLEY, A.M., MOWAT, G.N., MACASKILL, J.A. & HOLMES, P.H., 1982. Immunosuppression in bovine trypanosomiasis: Response of cattle infected with Trypanosoma congolense to foot-and-mouth disease vaccination and subsequent live virus challenge. Research in Veterinary Science, 32, 289–293.
- SNOW, W.F., WACHER, T.J. & RAWLINGS, P., 1997. Observations on the prevalence of trypanosomosis in small ruminants, equines and cattle, in relation to tsetse challenge, in The Gambia. Veterinary Parasitology, 66, 1–12.1
- SHERENI, W., 1990. Strategic and tactical developments in tsetse control in Zimbabwe (1981–1989). Insect Science and its Application, 11, 399– 409.
- SONES, K.R., NJOGU, A.R. & HOLMES, P.H., 1988. Assessment of sensitivity of Trypanosoma congolense to isometamidium: A comparison of tests using cattle and mice. Acta Tropica, 45, 153–164.
- SOULSBY, E.J.L., 1982. Helminths, Arthropods and Protozoa of Domesticated Animals. 7th edn. London: Bailliere Tindall.
- SPECHT, E.J.K., 1982. Seasonal incidence of helminths in sheep and goats in South Mozambique. Veterinary Parasitology, 11, 317–328.
- STEPHEN, L.E., 1966. Pig trypanosomiasis in tropical Africa. Commonwealth Bureau of Animal Health. Review series No. 8, Commonwealth Agricultural Bureau, Farnham House, Farnham Royal, Bucks., U.K.
- STEPHEN, L.E., 1970. Clinical manifestations of the trypanosomiases in livestock and other domestic animals. In: MULLIGAN, H.W. & POTTS, W.H., (eds). The African Trypanosomiases. London: George Allen and Unwin/ Ministry of Overseas Development.
- STEPHEN, L.E., 1986. Trypanosomiasis. A Veterinary Perspective. Oxford: Pergamon Press.
- STEPHEN, L.E. & MACKENZIE, C.P., 1959. Experimental Trypanosoma vivax infection in the horse. The Veterinary Record, 71, 527–531.
- SULIMAN, H.B. & FELDMAN, B.F., 1989. Pathogenesis and aetiology of anaemia in trypanosomaisis with special reference to T. brucei and T. evansi. Veterinary Bulletin, 59, 99–107.
- TABEL, H., 1979. Serum protein changes in bovine trypanosomiasis: A review. In: LOSOS, G. & CHOUINARD, A., (eds). Pathogenicity of Trypanosomes. Proceedings of a Workshop held at Nairobi, Kenya, 20–23 November 1978. Ottawa: International Development Research Centre.
- TAIT, A. & TURNER, C.M.R., 1990. Genetic exchange in Trypanosoma brucei. Parasitology Today, 6, 70–75.
- TAKKEN, W., TAYLOR-LEWIS, E.G. & WOODFORD, M.H., 1988. Field studies on animal trypanosomiasis in Mozambique. I. Effectiveness of the prophylactic drugs isometamidium chloride and pyrithidium bromide. Tropical Animal Health and Production, 20, 243–255.
- TEIXEIRA, A.R.L., 1987. The stercorarian trypanosomes. In: SOULSBY, E.J.L., (ed.). Immune Responses in Parasitic Infection, Immunology, Immunopathology and Immunoprophylaxis. Vol. III: Protozoa. Boca Raton, Florida: CRC Press Inc.
- THEILER, A., 1903. A new Trypanosoma, and the disease caused by it. Journal of Comparative Pathology, 16, 193.
- TOURÉ, S.M., 1971. Etude ultrastructurale des effets produits par diverses drogues trypanocides sur des trypanosomes. Revue d’Elevage et de Médecine vétérinaire des Pays tropicaux, 24, 381–392.
- TRAIL, J.C.M., D’IETEREN, G.D.M., COLARDELLE, C., MAILLE, J.C., ORDNER, G., SAUVEROCHE, B. & YANGARI, G., 1990. Evaluation of a field test for trypanotolerance in young N’Dama cattle. Acta Tropica, 48, 47–57.
- TRAIL, J.C.M., D’IETEREN, G.D.M., FERON, A., KAKIESE, O., MULUNGO, M. & PELO, M., 1990. Effect of trypanosome infection, control of parasitaemia and control of anaemia development on productivity of N’Dama cattle. Acta Tropica, 48, 37–45.
- TRAIL, J.C.M., D’IETEREN, G.D.M. & TEALE, A.J., 1989. Trypanotolerance and the value of conserving livestock genetic resources. Genome, 31, 805–812.
- TRAIL, J.C.M., MURRAY, M., SONES, K., JIBBO, J.M.C., DURKIN, J. & LIGHT, D., 1985. Boran cattle maintained by chemoprophylaxis under trypanosomiasis risk. Journal of Agricultural Science, Cambridge, 105, 147–166.
- TRUC, P., AERTS, D., MCNAMARA, J.J., CLAES, Y., ALLINGHAM, R., LE RAY, D. & GODFREY, D.G., 1992. Direct isolation in vitro of Trypanosoma brucei from man and other animals, and its potential value for the diagnosis of gambien trypanosomosis. Transactions of the Royal Society of Tropical Medicine and Hygiene, 86, 627–629.
- TURNER, C.M.R., BARRY, J.D. & VICKERMAN, K., 1988. Loss of variable antigen during transformation of Trypanosoma brucei rhodesiense from bloodstream to procyclic forms in the tsetse fly. Parasitology Research, 74, 507–511.
- URQUHART, G.M., 1980. The pathogenesis and immunology of African trypanosomiasis in domestic animals. Transactions of the Royal Society of Tropical Medicine and Hygiene, 74, 726–729.
- URQUHART, G.M. & HOLMES, P.H., 1987. African trypanosomiasis. In: SOULSBY, E.J.L., (ed.). Immune Responses in Parasitic Infections: Immunology, Immunopathology and Immunoprophylaxis. Vol. III. Protozoa. Boca Raton, Florida: CRC Press Inc.
- VAN DEN BOSSCHE, P., 1997. The control of Glossina morsitans morsitans (Diptera: Glossinidae) in a settled area in Petauke District (Eastern Province, Zambia) using odour-baited targets. Onderstepoort Journal of Veterinary Research, 64, 251–257.
- VAN DEN BOSSCHE, P., SHUMBA, W. & MAKHAMBERA, P., 2000. The distribution and epidemiology of bovine trypasonomosis in Malawi. Veterinary Parasitology, 88, 163–176.
- VAN DEN BOSSCHE, P. & DUCHATEAU, L., 1998. The effect of deltamethrin pour-on applied to cattle on the transmission of bovine trypanosomosis. Revue d’Elevage et de Médecine vétérinaire des Pays tropicaux, 51, 123–126.
- VAN DEN BOSSCHE, P. & MUDENGE, D., 1997. Prevalence of tsetse-transmitted trypanosomosis along the eastern/north eastern border of Zimbabwe. Zimbabwe Veterinary Journal, 28, 49–59.
- VAN DEN BOSSCHE, P., MUDENGE, D., MUBANGA, J. & NORVAL, A., 1999. The parasitological and serological prevalence of tsetse-transmitted bovine trypanosomosis in the eastern Caprivi (Caprivi District, Namibia). Onderstepoort Journal of Veterinary Research, 66, 103–111.
- VAN DEN BOSSCHE, P. & STAAK, C., 1997. The importance of cattle as a food source for Glossina morsitans morsitans Westwood (Diptera: Glossinidae) in Katete District, Eastern Province, Zambia. Acta Tropica, 65, 105–109.
- VALLI, V.E.O., FORSBERG, C.M. & MILLS, J.N., 1979. Pathology of T. congolense in calves. In: LOSOS, G. & CHOUINARD, A., (eds). Pathogenicity of Trypanosomes. Proceedings of a Workshop held at Nairobi, Kenya, 20–23 November 1978. Ottawa: International Development Research Centre.
- VICKERMAN, K., 1969. On the surface coat and flagellar adhesion in trypanosomes. Journal of Cell Science, 5, 163–193.
- VICKERMAN, K., 1985. Developmental cycles and biology of pathogenic trypanosomes. British Medical Bulletin, 41, 105–114.
- VICKERMAN, K. & PRESTON, T.M., 1976. Comparative cell biology of the kinetoplastid flagellates. In: LUMSDEN, W.H.R. & EVANS, D.A., (eds). Biology of the Kinetoplastida. Vol. I. London: Academic Press.
- VOLLER, A., BIDWELLL, D.E. & BARTLETT, A., 1975. A serological study on human T. rhodesiense infections using microscale ELISA. Tropenmedizin und Parasitologie, 26, 247–251.
- WACHER, T., RAWLINGS, P. & JEANNIN, P., 1988. Tsetse and trypanosomiasis in cattle: The interface of grazing patterns and its importance. In: Livestock Production in Tsetse Affected Areas of Africa. The AfricanTrypanotolerant Livestock Network. Proceedings of a Meeting. 23–27 November 1987, Nairobi, Kenya. ILCA/ILRAD 1988. Nairobi: English Press.
- WAINDI, E.N., GOMBE, S. & ODOUR-OKELO, D., 1986. Plasma testosterone in Trypanosoma congolense-infected Toggenburg goats. Archives of Andrology, 17, 9–17.
- WARNES, M.L., VAN DEN BOSSCHE, P., CHIHIYA, J., MUDENGE, D., ROBINSON, T.P., SHERENI, W. & CHADENGA, V., 1999. Efficacy of insecticide-treated cattle as a barrier to re-invasion of tsetse flies to cleared areas in north-eastern Zimbabwe. Medical and Veterinary Entomology, 13, 177–184.
- WEBSTER, P., 1989. Endocytosis by African trypanosomes. I. Three-dimensional structure of the endocytic organelles in Trypanosome brucei and T. congolense. European Journal of Cell Biology, 49, 295–302.
- WEBSTER, P., RUSSO, D.C.W. & BLACK, S.J., 1990. The interaction of Trypanosoma brucei with antibodies to variant surface glycoproteins. Journal of Cell Science, 96, 249–255.
- WEITZ, B.G.F., 1970. Hosts of Glossina. In: MULLIGAN, H.W. & POTTS, W.H., (eds). The African Trypanosomiasis. London: George Allen and Unwin/ Ministry of Overseas Development.
- WELBURN, S.C. & MAUDLIN, I., 1989. Lectin signalling of maturation of T. congolense infections in tsetse. Medical and Veterinary Entomology, 3, 141–145.
- WELLDE, B.T. & CHUMO, D.A., 1983. Persistence of Berenil in cattle. Tropical Animal Health and Production, 15, 149–150.
- WELLDE, B.T., CHUMO, D.A., ADOYO, M., KOVATCH, R.M., MWONGELA, G.N. & OPIYO, E.A., 1983. Haemorrhagic syndrome in cattle associated with Trypanosoma vivax infection. Tropical Animal Health and Production, 15, 95–102.
- WELLDE, B.T., CHUMO, D.A., REARDON, M.J., MWANGI, J. & ASENTI, A., 1989. Presenting features of Rhodesian sleeping sickness patients in the Lambwe Valley, Kenya. Annals of Tropical Medicine and Parasitology, 83, 73–89.
- WELLDE, B.T., HOCKMEYER, W.T., KOVATCH, R.M., BHOGAL, M.S. & DIGGS, C.L., 1981. Trypanosoma congolense: Natural and acquired resistance in the bovine. Experimental Parasitology, 52, 219–232.
- WELLDE, B.T., REARDON, M.J., CHUMO, D.A., KOVATCH, R.M., WAEMA, D., WYKOFF, D.E., MWANGI, J., BOYCE, W.L. & WILLIAMS, J.S., 1989. Cerebral trypanosomiasis in naturally infected cattle in the Lambwe Valley, South Nyanza, Kenya. Annals of Tropical Medicine and Parasitology, 83, 151–160.
- WELLS, E.A., 1972. The importance of mechanical transmission in the epidemiology of nagana: A review. Tropical Animal Health and Production, 4, 74–88.
- WELLS, E.A., 1976. Subgenus Megatrypanum. In: LUMSDEN, W.H.R. & EVANS, D.A., (eds). Biology of the Kinetoplastida. Vol. I. London: Academic Press.
- WENYON, C.M., 1926. Protozoology. A Manual for Medical Men, Veterinarians and Zoologists. Vol. I. London: Ballière, Tindall and Cox.
- WHITELAW, D.D., 1987. Late stage T. brucei infections and efficacy/ distribution of trypanocidal drugs. ILRAD Annual Scientific Report of the International Laboratory for Research on Animal Diseases. Nairobi: General Printers Ltd.
- WHITELAW, D.D., GARDINER, P.R. & MURRAY, M., 1988. Extravascular foci of Trypanosoma vivax in goats: The central nervous system and aqueous humor of the eye as potential sources of relapse infection after chemotherapy. Parasitology, 97, 51–61.
- WHITELAW, D.D., GAULT, E.A. & HOLMES, P.H., 1991. Development of an enzyme-linked immunosorbent assay for the detection and measurement of the trypanocidal drug isometamidium (Samorin) in cattle. Research in Veterinary Science, 50, 185–189.
- WHITELAW, D.D. & JORDT, T., 1985. Colestrol transfer of antibodies to Trypanosoma brucei in goats. Annales de la Société Belge de Médecine Tropicale, 65, 199–205.
- WHITESIDE, E.F., 1958. The control of animal trypanosomiasis in Kenya. A paper prepared for IACED Symposium 1958. Department of Veterinary Services, Kenya.
- WHITESIDE, E.F., 1962. Interactions between drugs, trypanosomes and cattle in the field. In: GOODWIN, L.G. & NIMMO-SMITH, R.H., (eds). Drugs, Parasites and Hosts. London: J. and A. Churchill.
- WHO, 1979. The African Trypanosomiases. Report of a joint WHO Expert Committee and FAO Expert Consultation. Technical Report Series 635. Geneva, World Health Organization.
- WILLET, K.C., 1970. Epizootiology of trypanosomiasis in livestock in East and Central Africa. In: MULLIGAN, H.W. & POTTS, W.H., (eds). The African Trypanosomiases. London: George Allen and Unwin/Ministry of Overseas Development.
- WILLIAMSON, J., 1970. Review of chemotherapeutic and chemoprophylactic agents. In: MULLIGAN, H.W. & POTTS, W.H., (eds). The African Trypanosomiases. London: George Allen and Unwin/Ministry of Overseas Development.
- WILLIAMSON, J., 1976. Chemotherapy of African trypanosomiasis. Tropical Diseases Bulletin, 73, 531–542.
- WILSON, A.J., 1969. Value of the indirect fluorescent antibody detection test as a serological aid to diagnosis of Glossina-transmitted bovine trypanosomosis. Tropical Animal Health and Production, 1, 89–95.
- WILSON, A.J. & CUNNINGHAM, M.P., 1971. Immunological aspects of bovine trypanosomiasis. IV. Patterns in the production of common antibodies. Tropical Animal Health and Production, 3, 133–139.
- WILSON, A.J., PARIS, J., LUCKINS, A.G., DAR, F.K. & GRAY, A.R., 1976. Observations on a herd of beef cattle maintained in a tsetse area. Tropical Animal Health and Production, 8, 1–12.
- WOO, P.T.K., 1970. The haematocrit centrifuge technique for the diagnosis of African trypanosomiasis. Acta Tropica, 27, 384–386.
- WOOLHOUSE, M.E.J., BEALBY, K., MCNAMARA, J.J. & SILUTONGWE, J., 1994. Trypanosome infections of the tsetse fly Glossina pallidipes in the Luangwa Valley, Zambia. International Journal for Parasitology, 24, 987–993.
- WOOLHOUSE, M.E.J. & HARGROVE, J.W., 1998. On the interpretation of age-prevalence curves for trypanosome infections of tsetse flies. Parasitology, 116, 149–156.
- WOOLHOUSE, M.E.J., HARGROVE, J.W. & MCNAMARA, J.J., 1993. Epidemiology of trypanosome infections of the tsetse fly Glossina pallidipes in the Zambezi Valley. Parasitology, 106, 479–485.
- ZWEYGARTH, E. & ROTTCHER, D., 1987. The occurrence of Trypanosoma (Nannomonas) simiae in the cerebrospinal fluid of domestic pigs. Parasitology Research, 73, 479–480.




