- Infectious Diseases of Livestock
- Part 3
- Bovine brucellosis
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Bovine brucellosis
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Bovine brucellosis (Brucella abortus infection)
Synonyms: Contagious abortion
Previous authors: J GODFROID, P P BOSMAN, S HERR AND G C BISHOP
Current authors:
J GODFROID - Professor, DVM, MSc, PhD, Faculty of Biosciences, Fisheries and Economics, UiT - The Arctic University of Norway, Hansine Hansens veg 18, Tromsø 9019, Norway and Faculty of Veterinary Science, University of Pretoria, Private, Bag X04, Onderstepoort, Gauteng, South Africa, 0081
R.L Santos - Professor Titular, DVN, PhD, Universidade Federal de Minas Gerais, Escola de Veterinária Departamento de Clínica e Cirurgia Veterinárias Av. Antonio Carlos, 6627 Belo Horizonte 31270-901, MG, Brazil
J. Guitian - Professor, LV, MSc, PhD, DipECVPH, FHEA, GradStat, Veterinary Epidemiology, Economics and Public Health, The Royal Veterinary College, University of London, Hawkshead Lane, North Mymms, Hatfield, Hertfordshire, AL9 7TA, United Kingdom
J.M. Blasco - Senior Researcher, DVM, PhD, Centro de Investigación y Tecnología Agroalimentaria de Aragón, Unidad de sanidad animal, Avda. Montañana 930, Zaragoza 50059, Spain
Introduction
Bovine brucellosis is a highly contagious disease caused mostly by Brucella abortus, a bacterium which occurs intracellularly in its mammalian host. Apart from causing characteristic mid- to late-term abortion and infertility in cows, B. abortus also occasionally causes orchitis and inflammation of the accessory sex glands in bulls. Other livestock and wild animal species (see Brucella infections in terrestrial wildlife), though of varying susceptibility, are sometimes infected.77 Bovine brucellosis is also an important zoonosis.8 In some countries, particularly in southern Europe and western Asia, where cattle are kept in close association with sheep or goats, infection and abortion can also be caused by Brucella melitensis.195 Occasionally, Brucella suis may cause an infection in cattle but has not been reported to cause abortion.69, 74
By visiting the OIE (World Organisation for Animal Health) website and using the WAHIS interface,153 information on the worldwide brucellosis status as well as those animal diseases listed by the OIE due to their implications for international trade or public health can be obtained. This information is regularly updated and is based on the emergency level of the situation and on monthly and annual reports sent to the Central Bureau of the OIE by national veterinary administrations and other official sources. In sub-Saharan Africa, brucellosis is an important disease in both humans and livestock. In general, the assessment of the occurrence of brucellosis is restricted to few published studies based on serological surveys and it is considered to be the highest in pastoral production systems.129 Recent studies show that brucellosis is highly prevalent among dairy herds in peri-urban areas of several sub-Saharan African countries.79, 139 The surveillance and control of brucellosis in sub-Saharan Africa is rarely implemented outside southern Africa. The rate of infection in humans is virtually unknown and public awareness is extremely low. Hence, the impact of brucellosis in terms of public health and social importance is rarely correctly addressed.129
It is suspected that bovine brucellosis was introduced into southern Africa with cattle imported from Europe,88 but there is also a possibility that it was introduced into the subcontinent much earlier during the migration of people and their cattle herds from other African countries.123 The first reliable record of its existence in South Africa was that of Gray in 1906 when he reported a serious outbreak of abortion among cattle near Johannesburg.88 Its presence was finally confirmed by Hall in 1913 when he isolated B. abortus from the stomach of an aborted bovine foetus.88 Outbreaks of abortion thought to be bovine brucellosis were first observed in Zimbabwe in 1906, and the presence of brucellosis was confirmed in that country in 1914. According to an obituary notice by Bevan in The Veterinary Record of 1957, Zimbabwe was the first country to show that B. abortus-infected cattle could transmit the pathogen to humans and that goats were not the source of the infection.7
The disease has a relatively high prevalence in southern Africa, especially in intensively farmed areas, and it is the most important bacterial cause of abortion on the subcontinent. It has an important economic impact on the beef and dairy cattle industries, especially as in 1990, 14,7 percent of the herds in South Africa were known to be infected and the losses to cattle farmers exceeded R300 million per annum.13
In recent decades, developing countries, particularly India, have increased their share in global dairy production. In India, brucellosis in cattle and water buffalo accounted for more than 95 percent of the total losses occurring due to brucellosis in livestock populations, and has been responsible for a median loss of US $ 3.4 billion in 2015.179 The impact of brucellosis and other zoonoses affecting livestock production has to be globally assessed by their impact on human health. Of note, worldwide, brucellosis is not ranked among the top diseases based on disability-adjusted life years (DALY) losses.129
Aetiology
Brucella abortus is a small, Gram-negative, non-sporulating, non-encapsulated coccus, coccobacillus or short rod, 0,6 to 1,5 µm in length and 0,5 to 0,7 µm in width.5, 50 The organism is not acid-fast but does resist decolourization by weak acids and thus stains red with Stamp’s modification of the Ziehl-Neelsen stain.5, 180
Most wild strains are fastidious and slow-growing, and require carbon dioxide (5 to 10 percent ) supplementation for primary isolation at an optimal growth temperature of 36 to 38 °C. Brucella abortus strain 19 is an attenuated strain of reduced virulence which is used for the production of a live vaccine (see Control). It grows well in a normal atmosphere at 37 °C.5 Complex media, containing serum, are required for the growth of B. abortus and, although most strains grow on sheep blood agar, the colonies may not be as distinctive as when grown on serum dextrose agar. Growth, on primary isolation, is seldom clearly visible before 48 hours of incubation, at which stage the colonies are usually 0,5 to 1,0 mm in diameter. The use of selective media, such as Farrell’s medium, may substantially enhance the chances of isolation by inhibiting the growth of contaminants.70 The growth rate of B. abortus may, however, be markedly retarded by selective media and for this reason such cultures should be incubated for five days or longer.5 Smooth colonies on a clear growth medium, such as serum-dextrose agar, are convex, entire-edged, have a smooth, shiny surface, and are pale yellowish-brown when viewed under transmitted light.5, 180 Considerable variation in colour and surface texture are found in the rough strains. Smooth forms are often markedly pathogenic whereas the rough variants are usually less so.
There is no single test by which B. abortus may be identified with absolute certainty, but a combination of growth characteristics, colonial and cellular morphology, staining properties, agglutinating antisera and biochemical reactions will allow an accurate identification.5 Eight biovars (biotypes) are recognized (biovars 1, 2, 3, 4, 5, 6, 7 and 9), which may be differentiated by phage typing, mono-specific antisera, biochemical reactions and growth inhibition tests.5 There are no proven differences in the pathogenicity of field strain biovars.142 Biovar 7 is reported to be a mixed culture and, because no authentic isolate of it has been made for many years, it is expected to be deleted from the list.5 Ninety percent of the isolates typed in South Africa are biovar 1 and 10 percent are biovar 2.25 In Zimbabwe, B. abortus biovar 1 has been isolated.128, 133
Recently, several molecular-based protocols have been developed for identification of Brucella spp. to the genus, species and biovar level,116, 163 whereas MALDI-TOF analysis is a promising approach for species identification.115
Epidemiology
Bovine brucellosis is dynamic disease, with emergence of new areas of infection and re-emergence of infection in areas where infection existed earlier. Some countries in the developed world (Australia, New Zealand, Canada, USA, Japan, the majority of European countries) are declared officially free from bovine brucellosis15 after implementing costly control and eradication programmes for decades.
In Latin America, brucellosis is one of the most important zoonoses because of its impact on public and animal health, and on the economy. Its distribution is closely associated with the distribution and density of livestock.119
Data extracted from a small number of quality-assured studies, indicate that brucellosis is endemic at high levels in both small and large ruminants in some Middle eastern countries, such as Egypt and Jordan.138
India is the world’s leading milk producer, contributing around 17 percent of the world’s total milk production and has the world’s largest dairy cattle population at around 300 million. In 2019, the overall true prevalence of brucellosis observed in cattle was 8.3 percent and in water buffalo 3.6 percent . The highest prevalence of brucellosis was reported in the state of Punjab in both cattle and buffalo (23.51 and 10.2 percent respectively).176
In the province of Punjab in Pakistan, a 5.6 percent seroprevalence was found in buffalo and cattle reared at 11 institutional-owned livestock farms.97
A recent systematic review indicated that the overall seroprevalence of brucellosis in dairy cattle herds in China was 1.9 percent during the period 2008-2018. In northern China, where the traditional agropastoral areas and the main animal breeding industry are located, the seroprevalence was >10 percent , whereas it reached only 5.5 percent in southern China.165
No accurate figures are available on the prevalence of brucellosis in cattle in Africa, as most reports are based on non-representative laboratory results.129 A series of cross-sectional studies carried out in peri-urban dairy production areas of seven countries in West and Central Africa have recently provided much needed prevalence estimates in these expanding production systems. Estimated herd seroprevalences were: Lomé (Togo) 62 percent, Bamako (Mali) 32.5 percent, Bujumbura (Burundi) 14.7 percent, Bamenda (Cameroon) 12.6 percent, Ouagadougou (Burkina Faso) 3 percent, Ngaoundere (Cameroon) 2.3 percent, Thiès (Senegal) 1.3 percent, Niamey (Niger) 1.2 percent, Dakar (Senegal) 0.2 percent and Niakhar (Senegal) <0.04 percent. The results of these studies confirm that Brucella spp. circulates among peri-urban dairy farms in the region with more than one third of the herds in some areas such as peri-urban Lomé or Bamako presenting evidence of infection. Interestingly, the studies provide some evidence suggesting that intensive systems that are gradually replacing the traditional practice of transhumance and in which cattle are kept confined all year round may increase the risk of infection.139
It is important to be reminded that an often forgotten shortcoming of brucellosis serology is the impossibility to infer which (smooth) Brucella sp. induced antibodies in the host.19, 79 In this respect, mixed farming and especially keeping small ruminants along with cattle, a common practice in low and middle income countries, is a recognized risk factor.79 The epidemiological situation seems to be very different between West and East Africa. Indeed, in West Africa, only B. abortus has been isolated from cattle and from small ruminants, 173 whereas in East Africa, cattle are infected with B. abortus, as indicated by studies in Tanzania127 and Uganda96, 135 or with both Brucella species, as recently described in Kenya.135 Lastly, recently, B. suis was isolated from cattle in Zimbabwe,111 whereas B. abortus, B. melitensis and B. suis have been isolated from cattle in Egypt.132
In South Africa, since 1968 when compulsory calfhood vaccination was introduced, the control scheme has made a considerable impact on the prevalence of the disease. The prevalence has decreased on a national scale from 10,5 percent in 1976 to 6 percent in 1979,27 1,9 percent in 1984/5,71 and 1,4 percent in 1988/9.11 The variation in prevalence is demonstrated by a survey of 90 percent of the dairy and beef herds in the Eastern Cape Province and Karoo between 1985 and 1989, which revealed a prevalence of less than 0,3 percent.98 The prevalence of brucellosis in about 5000 adult beef cows slaughtered at a large KwaZulu-Natal abattoir in 1981/2 was estimated to be less than 1,5 percent.26 Although these cows were drawn from all the provinces of South Africa, they originated mostly from KwaZulu-Natal.10 A recent study in cattle slaughtered at Gauteng abattoirs reported a 5,5% seroprevalence, while besides B. abortus, B. melitensis was isolated from tissue samples.107 This is the first documentation of B. melitensis in cattle in South Africa. The absence of brucellosis in cattle, goats and dogs in the Mnisi community in South Africa, bordering wildlife reserves including the Kruger National Park with multiple wildlife species infected with brucellosis, has been recently documented in a setting with vaccination, fencing and movement control.178
A retrospective sero-epidemiological survey of bovine brucellosis on commercial and communal farming systems in Namibia from 2004 to 2018 revealed that bovine brucellosis is endemic in Namibia at a low percentage primarily in communal cattle herds. Widespread vaccination of cattle and robust planned surveillance is recommended. Due to the low level of positive reactors, the risk of importing or exporting brucellosis through cattle was inferred to be minimal.125
A prevalence of 4,5 percent in dairy cattle and an overall prevalence of 1,4 percent in cattle herds tested in Zimbabwe have been reported, even though intensive management systems employed in Zimbabwe generally seem to favour the spread of bovine brucellosis.124 Cattle in the communal areas of Manicaland Province were free of brucellosis before 1985 while the prevalence was high in intensively managed herds in the Midlands and Mashonaland. However, with the establishment of dairy cooperatives in resettlement areas in Zimbabwe in 1980, cattle of unknown disease status were mixed and often grazed together on communal pastures. As a result of this practice the prevalence of brucellosis in one settlement scheme in Manicaland Province rose from 0,7 percent in 1986 to 3,3 percent in 1988. In Zimbabwe the prevalence of bovine brucellosis in the communal areas also varies from province to province. The prevalence is lowest in the Mashonaland province and highest in Manicaland and Matebeleland. The patchy distribution of brucellosis in the provinces (some herds are free of the disease, while others have a high prevalence) may be explained by the fact that some communal herds have been kept closed whereas other cattle owners have purchased infected animals to upgrade their stock.124 Brucella abortus and B. melitensis have been reported in several studies in animals in Zimbabwe but the extent of the disease remains poorly known.128 Importantly, the molecular characterization of strains showed that besides B. abortus, B. suis was isolated from cattle tissues.111
In Zambia, a seroprevalence rate of 20 percent of brucellosis in traditionally managed cattle was estimated in 2012, suggesting that control programmes are necessary for improved cattle productivity and reduced public health risk.136 The disease is reported to be widespread in Malawi,189 but its prevalence is unknown. In 2014, a study showed that little was known about brucellosis with very few animals tested.186 No information on brucellosis in livestock is available in the scientific literature for Lesotho, Eswatini (formerly Swaziland) and Botswana. In 2015, a study showed that 13/133 bovines in the Limpopo National Park in Mozambique tested positive by the Rose Bengal test. There is no other information in Mozambique.184
Cattle usually become infected after ingesting contaminated feed or water or licking an infected placenta, calf or foetus, or the genitalia of an infected cow soon after it has aborted or calved, at which time very large numbers of B. abortus are present, particularly in the placenta lochia.1, 142, 207 Animals may also become infected by inhaling organisms or through the conjunctiva.142 Calves may acquire infections in utero or they may become infected after ingesting infected colostrum or milk. Although most will rid themselves of the infection within a few months, a low percentage may remain infected for life142 and may spread the disease at their first and subsequent parturitions.9
Infected animals usually abort only once, and subsequent calves are carried to full-term although they may be infected. Approximately 2,5 to 9 percent of heifers born of seropositive cows may be latently infected but serologically negative until the middle of their first gestation or even later, when, for the first time, antibodies to B. abortus may be detectable or abortion may occur.9, 38, 63, 108, 109, 142, 202
There have been no controlled studies showing that bulls are more resistant to B. abortus than heifers and cows. Bulls may become infected in utero or during early calfhood by the oral route and retain the infection into adult life.166 In bulls, the testes and accessory sex glands may be affected and reveal inflammatory changes. Infected bulls may shed brucellae in their semen, seminal fluid and urine, and therefore in infected herds they should always be viewed with suspicion,160 particularly if artificial insemination using their semen is contemplated.126 The risk of introducing the disease into a herd through embryo transfer is probably negligible.35, 181
Brucella abortus is sensitive to pasteurization temperatures and its survival outside the host is largely dependent on environmental conditions. It may survive in an aborted foetus in the shade for up to eight months, for two to three months in wet soil, one to two months in dry soil, three to four months in faeces, and for eight months in liquid manure stored in tanks.9, 142 Generally, removal of infected animals from contaminated premises for one month is sufficient to prevent infection, provided the facilities have been properly disinfected.9
A contaminated environment or equipment used for milking or artificial insemination are further sources of infection. Permanent calving camps and lush pastures, particularly if they are wet and muddy, may play a very important role in the spread of the disease.55, 142
Large numbers of organisms are shed from the reproductive tract when infected cows abort. In those cows that lactate following abortion, milk, including colostrum, is an important source of infection, and bacteria may be excreted intermittently in milk throughout the lactation period. Urine and faeces of infected cattle are less important sources of the bacterium. The fluid in hygromas caused by B. abortus infection may contain large numbers of organisms but, because they are restricted to the lesion, they do not seem to be important in the spread of the disease.9, 64 There is a reduction in the numbers of organisms shed in the months following calving and abortion, and cows usually eventually become non-infective until the next pregnancy, when there is again a rapid increase of brucellae organisms in the reproductive tract.187 During subsequent pregnancies there is invasion of the gravid uterus and allantochorion,58 but abortion rarely recurs. Ninety percent of infected cows remain chronically infected; the infection may persist for life confined to the udder and target lymph nodes.142
Although B. abortus has been isolated from ixodid ticks and their eggs in Brazil, ticks probably do not play an important role in the transmission of the disease.83 The transmission of brucellosis by ticks, fleas or mosquitoes from an infected herd to a non-infected herd has never been proven.
Brucella abortus infection in sheep and goats may occasionally cause them to abort, but the infection does not spread in these species and they are apparently not a real danger to cattle unless there is close association between the species.120 Horses become infected particularly by ingestion of B. abortus-contaminated feed. In this species the organisms localize in bursae, tendons and joints and they are thus an unlikely source of infection for cattle.9
Several African wildlife species including African (Cape) buffalo (Syncerus caffer), hippopotamus (Hippopotamus amphibius), zebra (Equus burchelli), eland (Taurotragus oryx) and impala (Aepyceros melampus) — have tested serologically positive for brucellosis (see Brucella infections in terrestrial wildlife), but these species are probably not of great importance in the epidemiology of bovine brucellosis in southern Africa. This is possibly because of the relatively infrequent contact between cattle and wildlife.46, 47, 60, 102 There are few records of abortions in wildlife in southern Africa due to brucellae, although B. abortus biovar 1 has been isolated from the cotyledons of pregnant African buffalo at slaughter.85 Although serological surveys have revealed up to 23 percent positive reactors in African buffalo in the Kruger National Park in South Africa,12, 90 these animals probably do not constitute a significant source of infection for cattle because of the strict control measures to prevent the spread of foot and mouth disease across the boundaries of the park and from adjoining private nature reserves.24, 178 In the USA B. abortus has been isolated from bison (Bison bison) and elk or wapiti (Cervus elaphus) and in Italy from chamois (Rupicarpra rupicarpra) (see Brucella infections in terrestrial wildlife).
The typing of B. abortus isolates may yield important epidemiological information,73, 196, 200 as it allows sources of infection to be traced, particularly in countries where a number of biovars are present.57
In humans brucellosis or undulant fever caused, among other Brucella spp., by B. abortus is chiefly an occupational disease, occurring most often in veterinarians, stock inspectors, abattoir workers, laboratory personnel and farmers who become infected by contamination of abraded or intact skin or mucous membranes, or by inhalation during contact with infected cattle, foetuses or foetal membranes, and calves.9, 211 Humans may also become infected after ingesting unpasteurized dairy products containing field strains of B. abortus or after inadvertent self-inoculation with B. abortus strain 19 vaccine.9 After an incubation period of 5 to 30 days or longer, a mild, self-limiting or severe, prolonged disease may follow. Patients typically experience a fever that may be intermittent or irregular, hence the name undulant fever. Other common symptoms include chills, depression, weakness, headache, joint pain, generalized aches, sweating and fatigue, resembling the “chronic fatigue syndrome”. Enlargement of the liver, spleen and lymph nodes may occur, as well as signs referable to almost any other organ system but particularly the skeletal system.
Papular to pustular skin rashes that are sometimes evident on the arms of veterinarians following obstetric procedures have been attributed to allergy to brucellae, but sensitivity to other pathogens including Salmonella Dublin, Salmonella Typhimurium and Listeria monocytogenes, has also been incriminated.9
Pathogenesis
The establishment of infection is influenced by the size of the infective dose, virulence of the bacteria, and the resistance, age, sex and reproductive status of the animal.54, 142, 187
Brucella abortus readily penetrates mucous membranes such as those of the pharynx and alimentary tract, and survives and multiplies particularly in cells of the reticuloendothelial system.68, 187 After penetration, the organisms are phagocytosed by neutrophils and macrophages that carry them to the regional lymph nodes, where they multiply and induce lymphadenitis that may persist for months. Multiplication of the organisms here may be followed by bacteraemia, which may persist for several months, resolve itself, or be recurrent for at least two years in 5 to 10 percent of animals. Recurrence occurs particularly during pregnancy. During the bacteraemic phase, organisms are carried intracellularly in neutrophils and macrophages or free in the plasma and localize in various organs, especially the gravid uterus, udder and supramammary lymph nodes. Localization may also occur in other lymph nodes and the spleen, and in bulls in the testes and male accessory sex glands.28, 68, 142 Occasionally bacterial localization occurs in synovial structures, causing purulent tendovaginitis, arthritis or bursitis.64, 72, 99
Localization of the infection in the endometrium of the gravid uterus and in foetal membranes of cattle appears to be the result of the special affinity of the organism for erythritol,58, 103, 187, 203 elevated levels of which occur in the placenta and foetal fluids from about the fifth month of gestation. The chorionic epithelium becomes parasitized and infection extends to the placental stroma, blood vessels and ultimately, to the foetus.68, 187 There is considerable variation in the uterine and placental lesions in both natural and experimental B. abortus infections, and foetuses that become infected late in gestation may be aborted without any grossly recognizable placental lesions. Depending on the severity of the placentitis, abortion, premature birth or the birth of a viable or non-viable calf may result.68, 100, 142 The reasons underlying the genital tropism of Brucella and subsequent intense multiplication are still to be deciphered. It is important to note that, although erythritol is a Brucella preferential carbon source, the placenta and the male genital organs and fluids also have high concentrations of glycerol, lactate, and glutamate. It seems therefore plausible that there is a correlation between the presence and abundance of privileged nutrients in the male and female genital organs and the nutritional preferences of Brucella in some simple defined media.18, 112, 137, 212
The abundance of erythritol in the pregnant uterus results in the massive multiplication of Brucella organisms in this organ. The growth of most vaccine strain 19 B. abortus organisms, however, is generally inhibited by the presence of erythritol,67 but tolerance to erythritol by some strain 19 variants may be the cause of occasional persistent infections and abortions.52
In the pregnant animal brucellae replicate in the placental trophoblast during middle and late gestation after the cells have actively begun secreting steroids. The mechanism leading to abortion after mid-gestation in brucellosis is not known. Infected trophoblasts produce cortisol, a steroid hormone not produced in the non-infected placenta. This production, coupled with increased levels of oestrogen and prostaglandin synthesis and decreased production of progesterone, mimics the hormonal changes occurring at term in non-Brucella-infected cattle and leads to the initiation of parturition.68
Up to 35 percent of cows may be resistant to infection with B. abortus because their macrophages have a greater ability to kill B. abortus than that possessed by susceptible cows. The level of macrophage function, which is reduced in susceptible cows, plays a role in the establishment of chronic infections.87 This enhanced macrophage-killing activity is significantly greater in cows that are genetically resistant to infection, including that caused by Mycobacterium bovis, S. Dublin and S. Typhimurium as well as B. abortus.164 The bovine nramp1 gene, the homologue of the murine tuberculosis resistance gene, has been identified as a major candidate for controlling the in vivo resistant phenotype to Brucella infection. It has been demonstrated in a murine macrophage cell line transfected with the resistance- and susceptibility-associated alleles of the bovine nramp1 gene that these alleles severely affect the control and replication of B. abortus.20
Phagocytes have, on the one hand, developed antimicrobial defence mechanisms, such as oxidative burst, acidification of phagosomes, or fusion of phagosomes with lysosomes, to eliminate pathogens, while on the other, facultative intracellular bacteria have developed strategies counteracting the host cell defences, resulting in intramacrophagic survival. Recent studies have revealed that caveolae or lipid rafts anchored in the membrane of macrophages are implicated in the entry of brucellae into murine macrophages and mediate an endocytic pathway that avoids fusion with lysosomes.161 It has been shown that in human macrophages phagosomes rapidly acidify to a pH of 4,0 to 4,5 following B. suis infection and that this early acidification is crucial for intracellular replication, as neutralization results in bacterial elimination.161 In addition, if the phagosomal membrane is disrupted, then B. suis fails to multiply intracellularly.105 These results highlight that an intact, acidic phagosome called the ‘brucellosome’ is important for the intracellular trafficking of the Brucella-containing vacuole.104 A series of genes are involved in the adaptation of brucellae to three major stress conditions within the phagosome, i.e. acid stress, starvation and low oxygen tension.106 The Brucella intracellular pathogenesis, with emphasis on bacterial exploitation of the host endoplasmic reticulum-associated functions, and how autophagy-related processes contribute to the bacterium's intracellular cycle has been recently reviewed.39, 40
Long-term residence of brucellae in the phagosomal compartment of host macrophages is essential for their ability to produce disease in both natural and experimental hosts. The ability of Brucella spp. to interfere with intracellular trafficking of the Brucella-containing vacuole in both phagocytic and non-phagocytic host cells is highly dependent on a type IV secretion system encoded by the virB operon, which is conserved among Brucella species.101 Brucella infections inhibit spontaneously occurring apoptosis in human monocytes, thus preventing host cell elimination. This might represent a strategy for brucella development in infected hosts.86 Studies with Brucella mutants suggest that stationary-phase physiology is critical for their successful long-term residence in host macrophages,168 and reveals striking parallels between the strategies employed by rhizobiae to establish and maintain intracellular residence in their plant host and those used by the brucellae during their long-term survival in the phagosomal compartment of host macrophages.114
Cytokines such as IFN-γ, TNF-α, IL-2 and IL-12 control the intracellular growth of brucellae.18, 65, 84, 212, 213, 214, 215 Among these cytokines, the most important is IFN-γ, which strongly activates macrophages and induces enhanced intracellular killing of brucellae.18, 137, 212 In contrast to what is observed in murine macrophages, B. suis does not induce the production of TNF-α in human macrophages.37 Interleukin (Il) 10 favours Brucella infection by impairing a protective host immune response.208
In non-phagocytic cells, such as Hela epithelial cells, the Brucella bacterium initially interacts with compartments of the early endocytic cascade, then rapidly segregates from this intracellular pathway and associates with the autophagocytic cascade.159 During the late stages of infection, brucellae proliferate within the endoplasmic reticulum of host cells. They replicate extensively intracellularly61, 62, 159 without inducing obvious damage to the infected cell, and therefore seem to promote the survival of the cells for their own benefit.84 By studying Brucella infection in pregnant mice, it was discovered that B. abortus kills trophoblasts, which are important cells for maintaining pregnancy. This killing required an injected bacterial protein that triggered an endoplasmic reticulum stress response in the trophoblast.34 Eventually, in the bovine pregnant uterus, this extensive replication does lead to cell necrosis and acute inflammation, and to the release of huge numbers of bacterial cells from both the trophoblasts and foetal tissues.68
Clinical signs
The length of the incubation period of bovine brucellosis varies considerably. The incubation period has been variously defined, inter alia as the period between exposure and abortion.32 In bulls this period is even more imprecise as serological evidence of infection may be equivocal or lacking, and clinical signs may be absent. The length of the incubation period is also affected by the size of the infective dose, and the age, sex, stage of gestation, and immunity of the infected animal.55 In cows that do eventually abort, the usual length of the incubation period varies according to the time at which infection occurred. Cows infected at service abort after an average interval of 225 days, whereas those infected at seven months’ gestation abort about 50 days later.188 Congenitally infected calves may remain sero-negative for at least 18 months, after which they may manifest clinical signs. The longest recorded ‘incubation period’ in a cow is nine years.108
The abortion rate in infected herds is dependent on many factors and varies according to the susceptibility of the pregnant animals, management practices, the severity of the challenge, the period for which the herd has been infected, and various environmental factors such as the quality of pastures, which may affect cattle density, the climate and the topography.55, 58, 142 In fully susceptible herds, abortion rates vary from 30 to 70 percent . Caution should be taken when interpreting reported proportions of cows that abort following herd infection as this proportion should ideally be calculated as the ratio of cows that abort to cows that are pregnant and not to the total number of cows in the herd. The actual proportion of cows that are pregnant when the herd became infected is highly variable and often unknown. As a result, the ratio of abortions to total herd size, rather than to number of pregnant cows, is often reported. Increased public awareness, veterinary intervention, improved management practices and vaccination have all contributed to making the disease in these herds assume a more insidious, chronic form. In such herds, which are often closed, very few or no abortions occur and the disease is almost impossible to recognize clinically.55 Abortions typically occur at approximately five to seven months’ gestation, although some occur earlier or later. Weak, full-term calves that often die shortly after birth are sometimes encountered. About 20 percent of infected animals do not abort, while 80 percent of animals that abort as a result of B. abortus infection, do so only once.9 The placenta is not consistently retained after abortion but when it is retained metritis is common. Early abortion may result in a considerable reduction in the milk yield.59 Infection of the udder is clinically inapparent and the organ appears to be normal when palpated.131
In bulls, acute to chronic, uni- or bilateral orchitis, epididymitis, and seminal vesiculitis occasionally occur. The scrotal circumference may be normal or severely increased.27 Strain 19 vaccination may also cause orchitis.29
Uni- or bilateral hygromas (Figure 1), especially of the carpal joints, may be evident in some animals in chronically infected herds,67, 100 or may occasionally follow inoculation of heifers with strain 19.51 Progressive, erosive, nonsuppurative arthritis of the stifle joints has been reported in young cattle from brucellosis-free herds that were vaccinated with strain 19 vaccine.206
Pathology
Irrespective of the route of infection, the organism provokes regional lymphadenitis characterized by reticuloendothelial cell and lymphoid hyperplasia, as well as the infiltration of large numbers of mononuclear cells and some neutrophils, and a few eosinophils and plasma cells.100 Other lymph nodes in the body and the spleen may be affected later in the course of the infection but to a lesser degree.68

There is considerable variation in the severity of the uterine lesions at abortion. As the disease progresses lesions advance from acute (mild to severe) to chronic endometritis. Microscopically, the endometrium is infiltrated by lymphocytes and plasma cells, and some neutrophils. Microgranulomas may be scattered in the endometrium.68, 210
The most significant lesions after experimental infection at 6–7 months of gestation were necrotizing and suppurative placentitis and lymphohistiocytic mastitis in cows, and fibrinous pleuritis, fibrinous pericarditis and bronchopneumonia in aborted foetuses (premature expulsion of a non-viable foetus more than 15 days before the predicted date of delivery).207
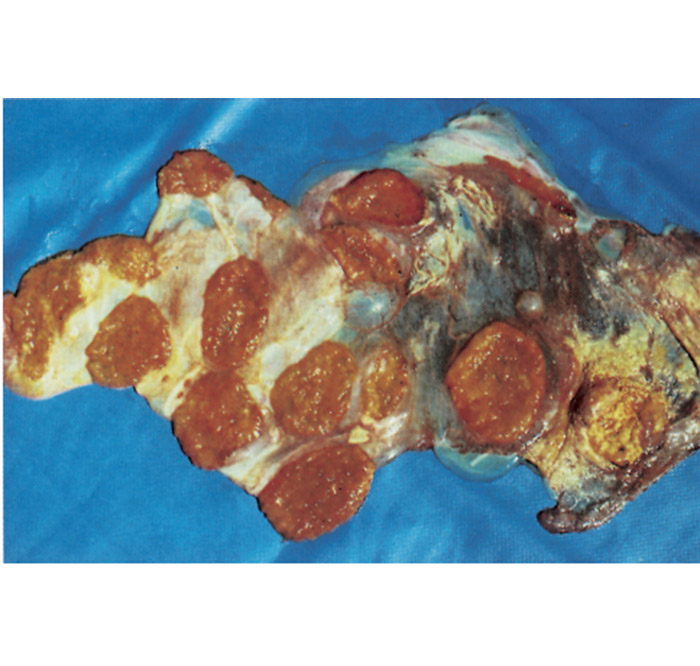
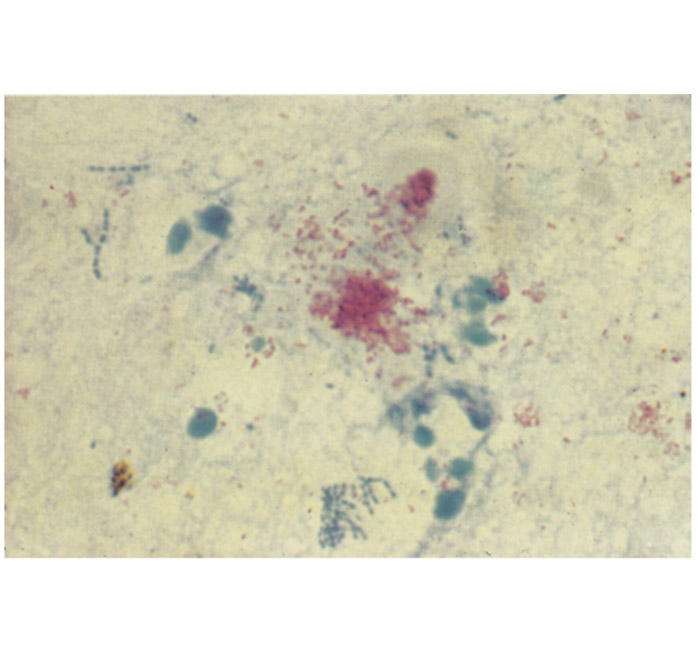
The chorion is not uniformly affected and large parts may appear quite normal. The lesions in and at the periphery of the cotyledons, as well as those in the intercotyledonary area vary in extent, appearing to be most severe adjacent to cotyledons. The affected cotyledons, or parts of them, are covered by a sticky, odourless, brownish exudate, and are yellowish-grey as a result of necrosis (Figure 2). Parts of the intercotelydonary placenta are thickened, oedematous, yellowish-grey and may contain exudate on the surface. Microscopically, the stroma of the chorion is infiltrated by numerous mononuclear cells and some neutrophils. Some chorionic villi are necrotic, while a fibrinopurulent exudate and desquamated necrotic chorionic epithelial cells are accumulated between the villi. Many of the chorionic epithelial cells are packed with numerous intracytoplasmic bacteria (Figure 3). Vasculitis, sometimes accompanied by thrombosis, may be evident in the chorion.68, 210
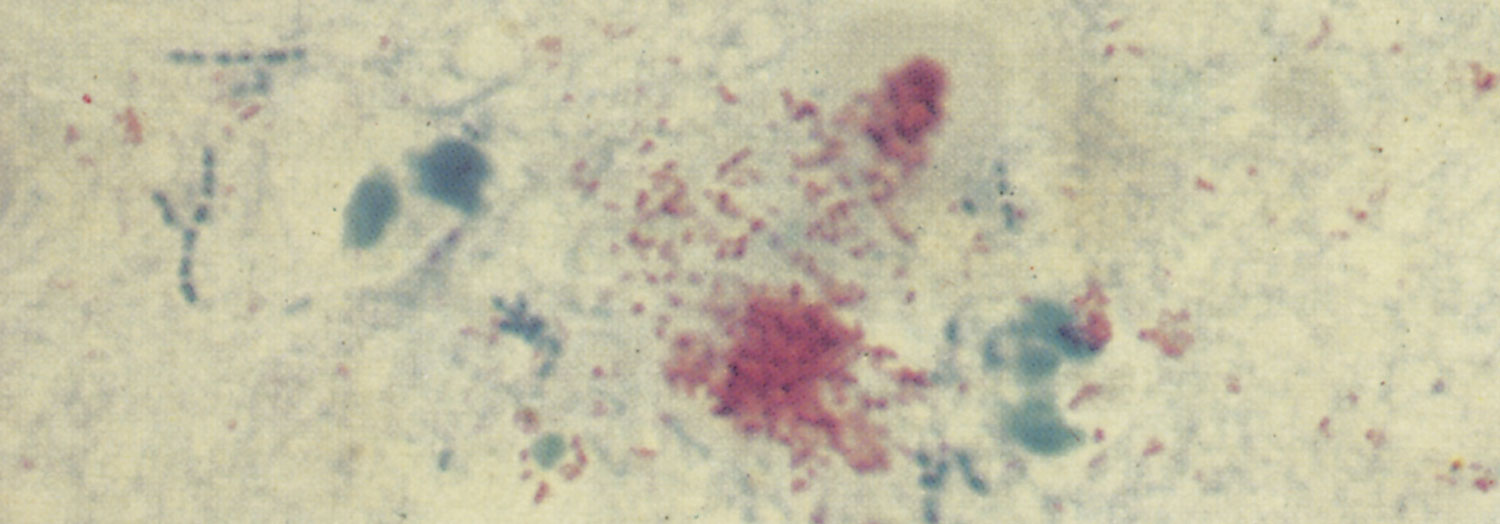
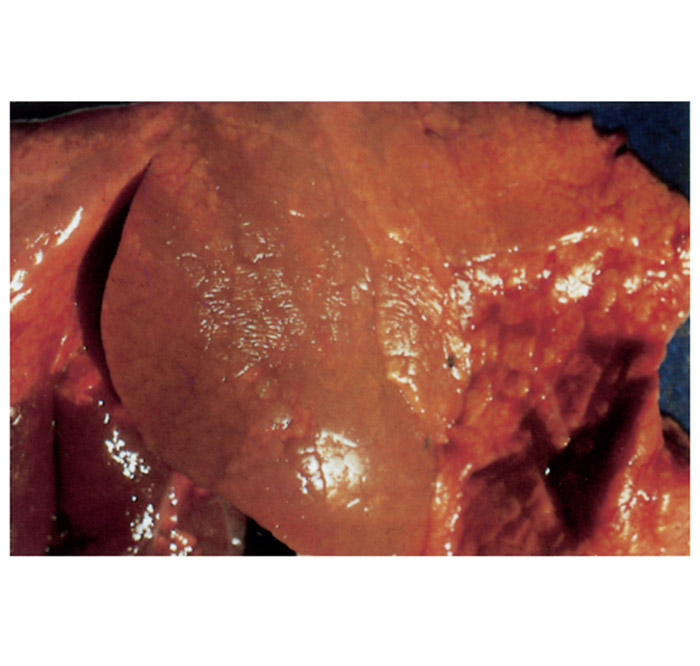
Some aborted foetuses have varying degrees of subcutaneous oedema and blood-tinged fluid in the thoracic and abdominal cavities, while the abomasal content is sometimes turbid, bright yellow and flaky. In some foetuses, greyish-white foci of pneumonia of 1mm or larger in diameter may be present, particularly in the apical lobes (Figure 4). Fibrinous pleuritis sometimes accompanies the pneumonia. The liver is usually enlarged, discoloured orange-brown and its surface may have a slightly uneven appearance. Many foetuses show no gross changes. Microscopically, most aborted foetuses reveal multifocal bronchopneumonia, characterized by the accumulation of cellular debris, neutrophils and macrophages in the lumen of the bronchi and bronchioli, patchy desquamation of bronchial epithelial cells, and mild to moderate infiltration of mononuclear cells and some neutrophils in the alveolar septa. Vasculitis of some of the pulmonary vessels may be seen. Isolated small foci of necrosis or microgranulomas are often found in the liver, but may also occur in the lymph nodes, spleen and kidneys. In most aborted foetuses it is not possible — or very difficult — to demonstrate organisms in tissue sections, notwithstanding that they may have been specially stained for brucellae.68 However, it is easy to demonstrate the organisms in smears made from the abomasal content or wall that have been stained with Stamp’s modification of the Ziehl Neelsen stain.5, 9
The udder in infected ruminants does not show any gross lesions, although the supramammary lymph nodes may be somewhat enlarged. 68, 210 Microscopically, infection of the udder is characterized by lymphoplasmacytic and histiocytic interstitial mastitis, while the regional lymph nodes show lymphoid hyperplasia, medullary plasmacytosis and sinus histiocytosis.68, 210
Acute orchitis is characterized by multifocal or diffuse necrosis of the testicular parenchyma, and focal, necrotizing epididymitis may occur. Microscopically the seminal epithelial cells are necrotic and desquamate; large numbers of organisms are present in them, while numerous leukocytes, particularly neutrophils, and fibrin occur in the affected tubuli and interstitial tissues. In the chronic stage, spermatic granulomas develop in the testicular parenchyma and epididymis in response to dead sperm.68, 210
In horses, infection may localize in the bursa between the nuchal ligament, the atlas and axis, causing poll evil. Fistulous withers caused by B. abortus is characterized by inflammation of the bursa between the nuchal ligament and the dorsal spines of the thoracic vertebrae. Chronic draining sinuses are formed in both conditions.55
Brucellae in cattle may localize in the carpal and other bursae and induce the formation of hygromas in which large numbers of the pathogen may be found.9, 71, 100
Diagnosis
In order to develop sensitive and specific diagnostic tests and more efficacious vaccines and to explore new therapeutic approaches in both human and animal brucellosis, it is important to assess both innate and acquired specific immune responses directed against brucellae. Because Brucella controls the traffic of its intracellular vacuole in phagocytes39, 40, 106 and in the trophoblast of the pregnant uterus,61, 62, 159 antibodies, while contributing to the protection against brucellae if present when infection occurs (for example after vaccination), are not as effective as that provided by cell-mediated immunity.
The humoral immune response is principally directed against the O-PS moiety of the smooth lipopolysaccharide (S-LPS).15, 148 The Brucella antiprotein antibody response is often delayed when compared to the anti-S-LPS response and is limited to animals that develop an active Brucella infection.113 Although there is evidence for a protective role of humoral antibodies directed either against LPS,43, 98 or outer membrane proteins,42, 98 the antibody response of cattle against B. abortus has been extensively used for the serological diagnosis of bovine brucellosis.5, 15, 148
Cellular immune responses contribute to the control of virulent and attenuated (e.g. vaccine strains) brucellae, more than the humoral response does.16, 17 It has been shown for many intracellular bacteria and protozoa that IFN-γ activates the microbial killing activity of macrophages.18, 134, 137, 212 Unfortunately, only few specific bacterial antigens involved in stimulating protective cellular immunity against Brucella are known.154 Murine macrophages infected with brucellae produce cytokines such as TNF-α and IL-12.214, 215 IL-12 seems to be critical in the mouse model because its depletion induces an exacerbation of the infection as well as an inhibition of the IFN-γ production.213, 215 IFN-γ is an important component of the type 1 cellular responses referred to nowadays as Th1 cell responses.134 These responses are principally characterized by the production of T1 cytokines, i.e. IFN-γ and IL-2, by CD4+ T helper cells and CD8+ T cytotoxic cells as well as IgG2a secretion by plasma cells.134 For extracellular bacteria or toxins, a Th2 cell response will be induced. This response is principally characterized by the production of Th2 cytokines, i.e. IL-4, IL-5 and IL-10, by CD4+ T helper cells as well as IgG1 and IgE secretion by plasma cells.134 According to the triggering mechanism, a precursor T helper cell will differentiate into either a Th1 or a Th2 cell.48 Live brucellosis vaccines are more effective than inactivated vaccines because the latter tend to induce a Th2 response.18, 213 Therefore, it is important to identify bacterial proteins that induce a Th1 mediated response, and it is critical to determine how the host first recognizes and then eliminates brucellae. However, although inducing a cellular immune response, few of these individual proteins induce host protection.42, 43
Current knowledge emanating from immunity studies in bovine brucellosis is incomplete, but parallels between the bovine and murine immune systems allow for some extrapolation of immunity to the disease in cattle.205
Other T lymphocyte subsets, such as γ δ T lymphocytes, may be important as mediator (secretion of IFN-γ and IL-2) or lytic effector cells in innate and acquired immunity. In humans, a particular subset of γ δ T cells impairs intracellular multiplication of B. suis in monocytes through soluble factor release and contact-dependent cytotoxic effects.65, 155 Bovine γ δ T cells from cattle infected with Mycobacterium bovis or vaccinated with a killed Leptospira borgpetersenii adjuvanted vaccine, recognize specific antigens and produce IFN-γ as part of an acquired immune response, which could play a role in protection.140, 167
Because of the variable incubation period and the often subclinical nature of the disease in most animals, a definitive diagnosis should be based on the isolation and identification of B. abortus and on positive serological results based on the detection of antibodies in blood, milk, whey, vaginal mucus, or seminal plasma.5, 9, 15
The isolation of B. abortus is important when dealing with specific groups or herds of animals when there is doubt about their infection status. Indeed, the Brucella species need to be identified in order to implement sound management measures.79 When a herd is known to be infected with B. abortus and where its environment is infected, serological reactors are viewed in a different light from those in a herd in which reactions might be due to the use of strain 19 vaccine. It is difficult to prescribe strict cut-off points when assessing serological results that will apply in all cases when strain 19 vaccine has been used. It is preferable to interpret serological test results according to the known infection status of the herd. As stated by Brinley Morgan:32
“I have found in my experience over the past twenty-five years working with Brucella spp. that you are invariably wearing at least two hats. One is where you are in a very clean environment where you can take an awful lot of chances, and the other hat is where you are working in an infected environment where you cannot afford to take any chances at all.”
Nevertheless, failure to isolate Brucella spp. in a herd may result in difficulty in interpreting serological results.
When detecting rare events, such as the occurrence of brucellosis at the end of an eradication programme, the emphasis is no longer put on intrinsic values of a test, but its positive predictive value, which relates to the clinical utility of the result.75 The aim of an eradication programme is not a ‘zero seropositivity’ situation, but the absence of infection, even if seropositivity is encountered. Indeed, the actual epidemiological situation (true incidence and prevalence rates) has to be taken into consideration: when tests are applied it must be borne in mind that the predictive values of tests vary according to the progress of the eradication programme and that criteria other than positive serology are important in an eradication programme.82
Diagnostic tests for bovine brucellosis are subdivided into three groups — tests for the demonstration of B. abortus organisms, those which detect immunoglobulins, and those dependent on allergic reactions to B. abortus.5, 9 The ‘ideal’ diagnostic test should detect infection early, during the long and variable incubation period; not be influenced by the presence of ‘non-specific’ antibodies; detect carriers; and differentiate between responses to vaccination and those due to field infection.15, 32, 148 There is no single available test that completely covers all these requirements.148
The tests described below are standard tests that are applied worldwide. Minimal requirements to ensure quality control of the reagents for the demonstration of B. abortus and the methods employed for the serological and allergic diagnosis of bovine brucellosis can be found in standard laboratory textbooks.5, 9, 15 A technical description of these tests can also be found in the OIE manual.15
Tests for the demonstration of B. abortus
Microscopic examination
Using Stamp’s modification of the Ziehl-Neelsen stain,180 B. abortus stains red against a blue background in tissue sections and smears (Figure 2). However, this colour reaction is not specific for Brucella spp., as Coxiella burnetii, Chlamydophila abortus and Nocardia spp. are also weakly acid-fast. Nocardia spp. can be differentiated from these organisms on morphological grounds, but it is difficult to differentiate C. abortus and C. burnetii from Brucella spp. beyond any doubt.5, 9, 15
Culture and typing
The specimens of choice include foetal membranes, lungs, stomach content, liver and spleen of aborted foetuses and full-term calves; and from live cows uterine discharge, milk or colostrum.5, 9, 15
The supramammary lymph node is the most suitable specimen from carcasses of adult animals, but the retropharyngeal, mandibular, iliac, or the prescapular and parotid lymph nodes may also be collected. Other specimens that may be submitted include uterus, milk, udder tissue, fluid aspirated from hygromas, male accessory sex glands and testes.53, 91, 142 Isolation may also be attempted from the semen or seminal plasma of infected bulls.55
The supramammary lymph nodes are preferred for isolation of B. abortus from animals that have been slaughtered and a 90 percent recovery rate from infected animals may be achieved.5 Recovery of B. abortus on culture from infected adult cows may approach 100 percent using the supramammary, parotid, mandibular and sub-iliac nodes.53 Isolation of the pathogen from infected heifers is best achieved by the culture of the mandibular lymph nodes, and this probably reflects the primary route of infection through the conjunctiva or the nasal or oral mucosa.53 When specimens of large groups of cows (five or more) must be cultured, collection of supramammary lymph nodes should be sufficient.67
In order to get valuable epidemiological information, isolated brucellae have to be typed (species and biovar) according to the standard biotyping methods described in textbooks. 5, 9, 15 Besides this method, molecular typing methods such as MLVA (Multiple Locus Variable-Number-Tandem-Repeat Analysis)110, 196 and Multi Locus Sequence Typing (MLST),201 as well as Whole Genome Sequencing (WGS), are nowadays used and have provided important information related to Brucella phylogeny, taxonomy and epidemiology. Representative B. abortus strains isolated worldwide could be subdivided by MLVA into three clades congruent with the clades B, C1, and C2 of MLST.196 Most sub-Saharan African strains were from West Africa and of clade B. They were predominantly isolated from cattle with some strains from dromedary camels, or humans. Clade B strains predominantly belonged to biovar 3.196
Polymerase chain reaction
Numerous polymerase chain reaction (PCR)-based assays have been developed for the identification of members of the genus Brucella.29, 59 Certain circumstances require the identification of the Brucella spp. involved. For epidemiological trace-back, strain-specific identification is helpful. Several strategies have been explored to differentiate between Brucella spp. and biovars, including locus specific multiplexing, e.g. PCR-RFLP at the omp2 locus44 and AMOS-PCR based on IS711 insertion sequence, allowing the differentiation of the vaccine strains 19 and RB51.30 A real-time PCR was developed in a multiplex format in 2004, allowing the rapid identification of Brucella spp., B. abortus, and B. melitensis in a single test.163 A multiplex PCR assay (Bruce-ladder) for rapid and simple one-step identification of Brucella has been developed and validated in 2008. It can identify and differentiate in a single step most Brucella spp. as well as the vaccine B. abortus strains S19 and RB51 and the vaccine B. melitensis strain Rev.1.116
Tests for the detection of specific immunoglobulins
The serological diagnosis of brucellosis began in 1897 with the development of an agglutination test by Wright.204 Problems such as positive serological reactions resulting from, for example, exposure to cross-reacting micro-organisms, were encountered. Improvements of existing tests and development of new serological tests have since then taken place.148 At present, several serological tests are used worldwide.15 The outer membrane of smooth Brucella spp. strains is composed of phospholipids, proteins and S-LPS. Most of the serological tests, particularly those using whole-cell suspensions as antigen, such as the slow (tube) agglutination test (SAT), the Rose Bengal test (RBT), the complement fixation test (CFT), most enzyme-linked immunosorbent assays (ELISA) and the milk ring test (MRT) have been developed to detect antibodies directed against the O-PS moiety of the S-LPS. 5, 9, 15
These tests are suitable for use in surveys, large-scale campaigns and in control/eradication programmes of the disease as well as for trading purposes.9, 15 Inexpensive, rapid and simple screening tests with high sensitivity followed by more specific confirmatory test(s) in case of positivity in the screening test, are used in control programmes. The ability of individual tests to correctly identify infected or non-infected animals (i.e. their sensitivity or specificity) can be enhanced by combining two or more tests and interpreting their results in parallel or in series, depending on whether the objective is to increase the negative or the positive predictive value. A systematic review of current immunological tests for the diagnosis of cattle brucellosis highlights the usefulness of some of the S-LPS based tests like RBT and ELISA.66 An epidemiological inquiry and a risk factor analysis also need to be undertaken.82
The worldwide use of strain 19 vaccine, which induces a persisting antibody response, led to the development of tests that could solve or at least reduce the problem of the interference of vaccination in order to differentiate vaccinated animals from infected ones.145, 148 This problem has led to the development of the RB51 vaccine that shows negligible interference in classical serological brucellosis tests.174 However, it is important to note that the insistence in recent literature on the lack of usefulness of all S-LPS or native hapten polysaccharide tests in areas where S19 vaccination is implemented is a misinterpretation that overlooks scientific and practical evidence.66
Reliance on serological tests alone for the diagnosis of brucellosis can be misleading and thus other tests, such as the brucellosis skin test, and a sound proficient epidemiological inquiry have to be implemented.121, 170
In acute bovine brucellosis an increase in the level of IgM in the serum is the first evidence of antibody response. IgG soon becomes the predominant antibody and usually persists for as long as the animal remains infected while the IgM levels wane. Of the two subclasses of IgG in the serum, IgG1 is the most abundant and predominant agglutinating and complement-fixing antibody.9, 149
In South Africa, serum samples are screened with the RBT, while the CFT is used as the confirmatory test. The SAT is sometimes used as a supplementary test and has some value in detecting IgM, the persistent and often predominant immunoglobulin resulting from vaccination with strain 19 vaccine.9, 32, 149 The MRT is also used to screen and monitor bulk milk samples.
Rose Bengal test
The RBT is a modification of the plate agglutination test. The antigen, which has been stained with Rose Bengal stain, is buffered at a pH of 3,65.5, 9, 15 At this level of activity, ‘non-specific’ agglutinins are destroyed and IgG, normally the most abundant antibody in the serum of infected animals, agglutinates strongly.9, 32 Equal volumes (30 µ) of test serum and antigen are mixed, shaken for four minutes and viewed over an X-ray viewer and any degree of agglutination is recorded as positive.6, 15
The test is inexpensive and easy to perform. False negative results are rare and are usually obtained during the more chronic stages of the disease. Despite improved specificity at an acid pH, a high percentage of false positive reactions occur usually due to the presence of IgM as a result of strain 19 vaccination.1 This test is prescribed for international trade in cattle by the OIE.15
Complement fixation test
This test is regarded throughout the world as being the confirmatory test for the serological detection of infected animals. It has been modified, standardized and adapted to a microtitre system.6, 15 Unlike the SAT, the titres do not wane as the disease becomes chronic.9, 170 Results are expressed in International Units (IU) and a cut-off point of 20 IU has been defined15 which is rigorously applied where strain 19 vaccine has not been used for several years, as is the case in most of the European Union (EU), the USA and Australia. Its strict application in countries enforcing compulsory strain 19 vaccination (such as South Africa) may often be problematic and sometimes leads to unacceptably large numbers of false positives, because vaccination induces serological titres. As a consequence, considerable expertise and experience are needed to certify herds or individual animals free of brucellosis when they are classified as positive by the test. As a rule, vaccine titres tend to decline faster than those due to infection with wild strains. The decline in titres is also dependent on the vaccine dose. Although the CFT is useful when differentiating calfhood vaccination titres from those due to wild-strain infections, there may be difficulty in distinguishing vaccine reactions from those caused by wild strains when animals have repeatedly been vaccinated (although calfhood vaccination alone is considered to be adequate) or after they have become sexually mature.23 Haphazard vaccination of heifers and vaccination of adult animals may result in much confusion in the interpretation of laboratory results, and it is therefore essential that accurate records of vaccination and birth dates be kept to allow correct interpretation of the results of the CFT. In the UK persistent reactors in brucellosis-free herds occurred in less than 0,5 percent of vaccinated heifers,32 but 16 percent of cows in brucellosis-free herds in New Zealand have been shown to develop CFT titres that persisted for at least 12 months after strain 19 vaccination.23 In South Africa, the use of low-dose vaccination of sexually mature heifers causes few persistent titres.26, 92 This test is prescribed for international trade in cattle by the OIE.15
Slow (tube) agglutination test
In a number of countries the SAT was and still is used as a screening test for eradication purposes. It is considered to lack specificity and some authors discourage its use, at least for trading purposes.148 Non-specific agglutination in sera is decreased by the addition of EDTA with no reduction in B. abortus agglutination titres of sera from infected cattle.76, 122 The SAT-EDTA is a very specific test and particularly useful in detecting new infections as early as two weeks after infection, as demonstrated in experimental conditions,82 but its usefulness in herds that are chronically infected is more limited because some infected animals will be classified as negative by this test because the infection is in a chronic phase.170 In South Africa, the SAT is still very usefully employed as a supplementary test for indicating the levels of serum IgM, the predominant immunoglobulin after vaccination with strain 19 vaccine.93, 94, 95
Indirect enzyme-linked immunoabsorbent assay
Indirect enzyme-linked immunoabsorbent assay (iELISA) is more sensitive for detecting antibodies to Brucella spp. than are the RBT, SAT and CFT, but great care must be exercised in animals vaccinated with strain 19 vaccine.36, 182 An iELISA has been developed and validated in South Africa. It has been suggested that this test could replace not only the currently used confirmatory CFT , but also the two in-use screening tests, namely the RBT and SAT.158 The iELISA has been used successfully throughout Europe in strategies aimed to substantiate and to maintain the status of ‘brucellosis-free countries’.81 When the herd prevalence is low, i.e., less than 1 percent , a pooled sera approach could be adopted by policymakers to reduce the cost of the surveillance programme and increase the number of samples being tested, as recently documented in Uruguay.22 iELISA can also be used on individual or bulk milk samples.192, 193 This test is prescribed for international trade in cattle by the OIE.15
Competitive enzyme-linked immunoabsorbent assay
The basis of this assay is the use of a selected monoclonal antibody (MAb) that competes with low affinity antibody. The competitive enzyme-linked immunoabsorbent assay (cELISA) using a MAb specific for one of the epitopes of the B.abortus O-PS has been shown to have higher specificity than the iELISA.151, 169, 183 The cELISA was reported to be able to eliminate cross-reaction problems in serological tests induced by strain 19 vaccination or infection with cross-reactive bacteria. Unfortunately, the cELISA only partially solves the problem. Indeed, persistent competing antibodies have been observed after Yersinia enterocolitica O:9 infections,82, 147, 152 and vaccination with strain 19.1 However, residual antibody activity due to vaccination or cross-reactive infection was less persistent than with the other tests.1 In a recent study, none of the cELISA kits studied could be recommended as a single screening test because of their low specificity.162 This assay is prescribed as an alternative test for international trade in cattle by the OIE.15
Fluorescence polarization assay
The fluorescent polarization assay (FPA) is a simple and rapid technique for measuring antigen/antibody interaction and may be performed in a laboratory setting or in the field.150 The mechanism of the assay is based on random rotation of molecules in solution. A fluorochrome-labelled antigen of small molecular weight (a fragment of the O-polysaccharide (O-PS) of B. abortus S-LPS, for example), is added to serum or other fluids to be tested. If antibody is present, attachment to the labelled antigen will cause its rotational rate to decrease and this decrease can be measured. The FPA has received very promising reports.162 This test is prescribed as an alternative test for international trade in cattle by the OIE.15
Milk ring test
The milk ring test (MRT) is used to detect antibodies in milk. The development of a positive reaction is dependent on two reactions: (i) fat globules in the milk are aggregated by milk antibodies (fat-globule agglutinins); and (ii) stained Brucella cells (antigen), which are added to the milk, are agglutinated by the Brucella antibody/fat globule complexes, which rise to form a coloured cream layer at the top.5, 9, 15 This is a sensitive screening test used on bulk milk samples either to detect infected animals on a herd basis or to monitor clean herds. It should be carried out at least quarterly to ensure that all animals that come into lactation are screened. The sensitivity of the MRT is somewhat reduced when it is applied to large herds with few reactors but this loss in sensitivity in large herds of 150 or more animals can be counteracted to some extent by decreasing the ratio of antigen to milk in the test.5 Despite its reduced sensitivity in large herds, the MRT has been very successfully used to monitor the brucellosis-free status of dairy herds.9, 55 In the EU, after a positive MRT performed on a bulk milk sample has been obtained, the cows that had supplied the milk are individually tested by serology in order to detect those that are infected.81
Factors that may cause false positive results include a high prevalence of mastitis; a high proportion of cows in early or late lactation; recent (within three to four months) vaccination with strain 19 vaccine; and souring of milk. Milk samples may be preserved for testing by adding 0,5 ml of a formalin solution (prepared by mixing 7,5 ml of 37 percent formaldehyde with one litre of distilled water) to a 10ml milk sample. The duration and temperature at which samples are stored (in particular excessive heating such as storing for longer than five minutes at 45 °C) may cause false negatives. Pasteurized milk cannot be effectively tested by the MRT.5
Several countries have replaced the MRT by a milk iELISA. Although this latter technique has not been standardized, it is prescribed for international trade in cattle by the OIE.15
Test to demonstrate an allergic reaction to B. abortus
A skin test for the diagnosis of brucellosis has been used in extensively managed herds in New Zealand.121 The sensitivity of the test was considered to be low at the animal level and its specificity exceeded 99 percent , and thus it was claimed that this test is a useful low-cost method of identifying infected herds rather than individual animals.5 This test has recently been re-evaluated in the EU because of the emergence of ‘False Positive Serological Reactions’ (FPSR), i.e. non-specific reactions in all serological tests, that have emerged throughout Europe.82 These have been documented in Belgium and France since 1990, affecting up to 15 percent of the herds tested in some regions that are free of brucellosis. Yersinia enterocolitica O:9 infections have been shown to be responsible for these FPSR.82, 199 Experimental studies have shown that the brucellosis skin test is the only test that is able to discriminate between Y. enterocolitica O:9 and B. abortus infections beyond any doubt.82, 170 It is now a recommended herd test for brucellosis by the OIE15 and as an official test in the EU (Directive 64/432), particularly when monitoring is made difficult due to non-specific brucellosis serological reactions.170 However, calfhood vaccination using strain 19 may complicate interpretation of the brucellin test by inducing prolonged sensitivity to brucellae allergens.170
A diagnosis of bovine brucellosis based on in vitro antigen-specific IFN-γ production, which can be regarded as an in vitro correlate of the brucellosis skin test, has been developed.198 Unfortunately this test has been shown to be less specific than the brucellosis skin test. 82
Differential diagnosis
Numerous infectious agents may cause foetal loss and abortion in cattle. A multidisciplinary approach in terms of diagnostic tests is necessary to make a definitive diagnosis.89 Microscopic examination of smears or histopathological sections, particularly of the placenta, stained by the modified Ziehl-Neelsen method, may present difficulties in differentiating B. abortus morphologically from Coxiella burnetii or Chlamydophila abortus. Brucella abortus can be distinguished from C. burnetii and C. abortus by PCR, fluorescent antibody techniques and serological tests.50 Apart from these agents there are numerous viral, bacterial and parasitic causes of abortion in cattle that should be considered.
Brucella abortus may cross-react serologically with Escherichia coli serogroup 0:157, Y. enterocolitica serovar O:9, Salmonella serotypes of the Kaufmann-White group N, Francisella tularensis, Pseudomonas maltophilia, and Vibrio cholera49 because the immunodominant O-chain of S-LPS of these bacteria contains antigenic motives (epitopes) that may be detected in serological tests for brucellosis using whole B. abortus cells or S-LPS extracts.197 Such FPSR induced by these organisms are probably not of great significance in the early phase of eradication campaigns but when the prevalence of the disease has been reduced to a very low level, then this phenomenon may jeopardize the success of the eradication programme.82
Control
Treatment
Cattle suffering from bovine brucellosis are generally not treated. Brucella spp. may undergo L-transformation when exposed to certain antibiotics, such as penicillin and oxytetracycline.9, 19 The effect that such cell wall deficient forms have in preventing serological detection and the resultant creation of carrier animals requires further investigation.19
Vaccination
In 1906, Bang observed that cattle could be protected against brucellosis by vaccinating them with live cultures. Since then three strains of B. abortus have been used for the preparation of vaccines and have been studied extensively: strain 19, a smooth strain, used as a live attenuated vaccine;33, 145 strain 45/20, as a rough killed vaccine;123 and, more recently, strain RB51, as a rough live attenuated vaccine.174
The minimal requirements for vaccine production have to be followed and each batch of live vaccine must conform to the minimum standards set by the OIE.15 These include viability, pathogenicity and ability to immunize guinea pigs and/or mice against challenge with a virulent strain of B. abortus.15
Strain 19 vaccine
Because of its relative safety, potency, practicality of production and convenience of use in cattle, strain 19 remains the most acceptable and the most widely used vaccine against bovine brucellosis.144, 145 It was first described by Buck in 1930. After being kept one year in the laboratory after primary isolation from the milk of a Jersey cow, the strain was attenuated in guinea pigs.33
Strain 19 differs from other B. abortus biovar 1 strains in its requirement for carbon dioxide and sensitivity to thionine blue, penicillin and Safranin O. It is the only Brucella sp. strain that is inhibited by erythritol. 5, 9, 15 The mutation in the erythritol catabolism genes has been determined.172 Vaccination with strain 19 vaccine increases resistance to B. abortus but does not induce absolute immunity, and vaccination with it is not curative, i.e. if an animal is infected, vaccination will not cure the infection.143 The increase in resistance following vaccination has been termed ‘relative immunity’ since it is estimated to be only about 70 percent effective against field challenge by preventing unrestricted multiplication of B. abortus in the uterus and mammary glands.9, 144, 145
The main disadvantage of strain 19 vaccination is the induction of post-vaccinal antibodies that are detected in serological tests. At present, there is no single individual test that can be used to discriminate between antibodies induced by vaccination and those induced by infection, although newer tests (or combinations of tests) have been developed to reduce this problem.148
Strain 19 vaccine must be stored correctly and the cold chain maintained to retain its full potency. Lyophilization, however, has proved successful in the preservation of the vaccine.9 On reconstitution, lyophilized vaccine should be used on the same day, preferably within three hours.
Strain 19 and RB51 are the only brucellosis vaccines currently allowed for use in South Africa. Traditionally, the dose of strain 19 vaccine administered subcutaneously to heifer calves at four to eight months of age contains 5 × 1010 viable Brucella cells.9, 15, 144 The calfhood strain 19 vaccine produced by Onderstepoort Biological Products in South Africa contains 4–12 × 1010 viable cells per dose, and a single dose of this vaccine administered to heifers at the age of five months generally provides life-long relative immunity, although virtually all of them will have lost their serum antibody titres by the age of 16 to 18 months.2, 15, 144 The vaccination of heifers practically eliminates the occurrence of abortions in a herd.144 A gradual reduction in the prevalence of bovine brucellosis may be expected when 80 percent of the female population has been vaccinated and this vaccination pressure (‘herd immunity’) must be maintained for optimal results.144, 145
A policy of using a reduced-dose vaccine containing 3 × 108–9 organisms/dose in heifers of 4 to 12 months of age has been shown to provide the same degree of protection as the classical dose, and has therefore been the only vaccine available in the USA and in Europe.9, 15, 21, 144 Its use in the USA, however, has been prohibited since 1996 when the RB51 vaccine was approved for use.174
Administration of strain 19 vaccine into the conjunctival sac (one or two doses of 5–10 × 1010 at 4 and 8 months of age, respectively) results in good protection and the almost complete elimination of serological reactions in heifers and adults.144, 146, 202
In the event of an outbreak, the vaccination of adult cattle with strain 19 vaccine may be advantageous, particularly in large dairy herds and in herds where a large proportion of the animals have not previously been vaccinated.123 116 Mature cows inoculated with a 3–10 × 108 organisms/dose are protected for at least 12 months, although most (90 to 95 percent ) of the vaccinated animals lose their CFT titres within six months. Five to ten percent of animals may, however, remain serologically positive for 8 to 12 months or longer and strain 19 organisms may be isolated from their milk.3, 17, 67 It has been shown that the culling of infected cows and the stage of gestation at vaccination will affect the efficacy of strain 19.54, 56 The disadvantages of vaccinating adult cows include the development of vaccine reactions, a positive MRT, and the stigma attached to vaccinated adult animals because of their association with positive herds.145
In South Africa, adult vaccination is restricted by law to individual farms and may only be given when written permission by the State to do so has been granted. However, this practice may complicate the interpretation of serological results for a considerable period. Recently, a study documented the persistence of antibodies in a South African communal farming setting for over a year to 4,5 years after 'high' dose S19 vaccination, which can be difficult to differentiate from a response to infection with wild-type B. abortus.177
In order to be able to differentiate infected animals from vaccinated ones, besides use of the CFT, the following epidemiological criteria are taken into account:
- the herd is closed and has been closed for at least two years;
- the herd is closed except for introductions from herds certified free of brucellosis;
- there is no evidence of spread of infection within the herd;
- all cows that aborted were bacteriologically negative for B. abortus;
- the lymph nodes of animals with high titres, and which have been slaughtered, are negative for wild-type strains of B. abortus and/or have yielded strain 19 on culture;
- the foetal membranes and uterine discharges of cows exhibiting high titres are negative on culture;
- milk samples from reactors yielded strain 19 and/or are negative for the wild-type strain of B. abortus; and
- the possibility of infection having been introduced from a neighbouring infected herd or infected wildlife is ruled out.
Strain 19 vaccination may occasionally cause orchitis in bulls.27, 190 Because it has also been found in the semen of vaccinated bulls, its use in males has been restricted or prohibited.145 Uni- or bilateral hygromas, especially of the carpal joints, may occasionally follow inoculation of heifers with strain 19 vaccine.206 Progressive, erosive, non-suppurative arthritis of the stifle joints has been reported in young cattle in brucellosis-free herds that had been vaccinated with strain 19 vaccine.51 Systemic reactions following vaccination are rare and temperature reactions varying from 40,5 to 42 °C, lasting for two to three days, may occur in addition to swelling at the injection site.27 Abortion in less than 1 percent of vaccinated cows in late pregnancy has also been reported.145 There is no evidence of the spread of strain 19 to unvaccinated cattle, although the organism can be excreted in milk for two to three months in vaccinated cows after calving or abortion.145 Erythritol- tolerant isolates of strain 19 have been isolated from milk and aborted foetuses in New Zealand and South Africa.23, 26
Although strain 19 organisms may be intermittently present in the milk of vaccinated cows, there is no evidence of infection in humans who have ingested milk containing them. However, undulant fever in humans has been reported after accidentally being injected with the vaccine.9, 211
Strain 45/20 vaccine
In order to avoid post-vaccinal antibodies, rough vaccine strains have been developed and tested. Brucella abortus smooth strain 45/0 was isolated in 1922, and after 20 passages in guinea pigs a rough derivative named strain 45/20 was obtained.130 Strain 45/20, when first used as a live vaccine, was inclined to lose its rough characteristics and revert to the smooth form and therefore it has only been further used as a killed vaccine.185 Strain 45/20 has been used with success in an inactivated vaccine incorporating an oil adjuvant,57 however, this is not as protective as strain 19 vaccine in animals inoculated when less than nine months of age. Two consecutive vaccinations 6 to 12 months apart with strain 45/20 vaccine are usually recommended. Large unsightly granulomas may develop at the site of injection following inoculation of some batches of strain 45/20.58 This vaccine is no longer used.175
Strain RB51 vaccine
Since 1996, B. abortus rough strain RB51 has been the official vaccine used in the USA for the prevention of brucellosis in cattle.147 Certain other countries, mainly in Latin America, have officially approved the vaccine, but their vaccination protocols differ.145, 171 In 2002 Spain and Portugal received limited approval to conduct field trials. RB51 was approved for use in South Africa in 2002.
This vaccine strain is a rough rifampicin-resistant mutant of B. abortus strain 2308, a smooth virulent B. abortus biovar 1 strain.174 It is very stable and no reversion to smoothness has been described in vivo or in vitro. The genetic mutations that are responsible for the roughness and the attenuation of strain RB51 have been identified in the wboA gene encoding a glycosyltransferase, an enzyme essential for the synthesis of O antigen.77, 194
The organism behaves as an attenuated strain in a variety of animals including guinea pigs, in which it is rapidly cleared from the tissues and does not induce abortions. It is efficient in preventing Brucella infections in mice.175 Although producing low levels of O-chain,45 strain RB51 does not usually induce the production of O-chain antibodies in cattle that can be detected in the classical brucellosis serological tests regardless of their age, the dose they received and the frequency of injections.175
More than five million heifers have been vaccinated subcutaneously with a dose of 1–3,4 × 1010 cells without noticeable side-effects.175 This vaccine has been found to have reduced abortifacient effects in cows, but placentitis was reported when used at a full dose.156, 191 However, pregnant cows can usually be safely vaccinated with a reduced dose of 109 cells.157
Few experimental studies have been conducted in cattle to compare the vaccine potency of strain RB51 with that of strain 19. Strain RB51 vaccine has not been reported to induce a higher degree of protection than strain 19 vaccine.41 Although, under field conditions, strain RB51 vaccine has been reported to be superior to strain 19 vaccine,118 more information is needed in order to assess the intrinsic value of different strain RB51 vaccination protocols in cattle. An advantage of RB51 vaccination is that antibodies induced by it are not detected by the currently prescribed serological tests for brucellosis. Non-pregnant adult animals can therefore be vaccinated annually, thus boosting the individual and herd immunity.
Importantly, three human cases of brucellosis attributed to strain RB51 have been reported in the USA for the period 2017-2019. Each case was associated with consumption of domestically acquired unpasteurized (raw) milk. RB51 infection is missed by tests normally used for diagnosis. In addition, RB51 is resistant to rifampin, a first-line antibiotic used to treat human brucellosis. Therefore, clinicians should consider RB51 infection and inquire about raw milk consumption as part of the patient’s exposure history.141
BS2 vaccine
A smooth strain of B. suis, biovar 1 (strain 2) has been in use as an oral vaccine to control brucellosis in cattle, sheep, goats and pigs in China since 1971.209 This vaccine protects cattle against B. abortus, is safe if administered orally, does not induce persistent antibody titres and can be administered without having to restrain the animal.209 Although it has been used widely in China, its use is not recommended by the OIE.15
Management of control programmes
The planning and management of control or eradication programmes for brucellosis have been investigated in many countries, and by the World Health Organization.9 In the EU, a task force for monitoring disease eradication in the member states was created in 2000 with the objective, as far as bovine brucellosis is concerned, of expanding eradication schemes and improving the cost-benefit ratio of eradication programmes co-financed by the EU (Council Decision 90/ 424/EEC).81 Strategies based on the prevention of the spread of the disease between animals, the monitoring of brucellosis-free herds and zones, the elimination of infected animals by test and slaughter, strict ‘one-way’ movement of infected and suspect cattle, mass immunization to reduce the infection rate, and supporting specific education and training programmes, have all received attention.9, 55
In South Africa, bovine brucellosis is listed as a State-controlled disease, and its control and eradication are given high priority by the Directorate of Animal Health. The control and eradication of bovine brucellosis in South Africa is broadly based on a test-and-slaughter policy, the vaccination of heifer calves, and the prohibition of the sale and movement of infected cattle. All sera are first screened with the RBT and those that are negative are usually not tested further. All sera that test positive are subjected to the CFT. In order for a herd to qualify for accreditation in the official South African Bovine Brucellosis Scheme, all animals must be CFT-negative in the final two tests. Herds with a prevalence of 10 to 20 percent have been cleared of the disease while incurring losses that were still economically acceptable.92 Good progress towards achieving eradication has been made in some regions in South Africa; in 1975 the prevalence in KwaZulu-Natal was estimated at 9,0 percent and this was reduced to 0,6 percent by 1988.11 However, bovine brucellosis has not been eradicated in South Africa. No recent country-wide seroprevalence estimates are currently available. A recent study in cattle slaughtered at Gauteng abattoirs reported a 5,5 percent seroprevalence,107 while the absence of brucellosis in livestock in the Mnisi community bordering the Kruger National Park has been documented in a setting with vaccination, fencing and movement control.178
It is not appropriate to be prescriptive when recommending a strategy for the control or eradication of brucellosis that will be applicable or practical in all circumstances. Broadly based national strategies and policies will invariably differ in time according to the situation in each instance. The approaches to be adopted when attempting to eradicate the disease will vary markedly according to whether one is dealing with an infected herd or an uninfected (vaccine reactions) herd. Control measures on individual farms are also frequently affected by a variety of other factors. These include the infection rate in the herd, duration of infection, the abortion rate, whether an extensive or intensive farming system is being practised, rainfall, whether it is a beef or dairy herd, vaccine cover, the availability of, or necessity for, replacement stock, the financial situation of the producer, his proximity to a veterinary laboratory, neighbourhood cooperation, milk and slaughter quotas, slaughter prices of dairy cows, the availability of camps and suitable calving facilities, availability of veterinary guidance, and the handling facilities necessary to collect blood samples from large groups of cattle.9, 55
The following principles have been found to be crucial to the successful control or eradication of brucellosis:55
- reliable veterinary services,
- reliable diagnostic techniques,
- close cooperation and joint effort by the owner, laboratory, veterinary services and veterinarian,
- good records,
- education of the farming community and public awareness,
- established procedures for handling the disease,
- controlled movement of cattle, and
- permanent marking of infected and vaccinated animals.
Vaccination in conjunction with other measures can be used effectively in the control or eradication of brucellosis and, in general, it is recommended that all heifer calves be immunized where the prevalence of brucellosis in an area, region, province or country is high, and that cessation of immunization with strain 19 vaccine only be considered when the prevalence of the infection is reduced to 0,2 percent or less.9 At that time, a test-and-slaughter, management and hygiene programme should be implemented.
Brucella abortus remains endemic in low-resource settings in which an intensive vaccination programme that reaches optimal coverage may not be realistic in the short term. In fact, it has been suggested that in some of these settings the prevalence of infection may be increasing as more intensive livestock systems emerge.139 Given the potential economic and public health impact of infection, it is important that progressive control strategies (combining livestock vaccination, gradual culling and environmental sanitation) that can be sustained with limited resources are considered.
The reduction in numbers and the rapid elimination of all infected animals are essential in the control or eradication of brucellosis.9 All calves in infected herds should be regarded as potential sources of infection. The ‘two-year breakdown syndrome’ (that is those previously infected herds which have apparently been clear for about two years and in which a ‘breakdown’ occurs after this period) has been ascribed to latently infected calves that remain serologically negative until at least mid-gestation or later during their first pregnancy.58
Cows should calve in an environment that can be thoroughly cleaned and disinfected with 2,5 percent formalin. Wet and well-grassed calving camps should be avoided, and vehicles used for transporting infected animals should be disinfected after use. Isolation of cows prior to and after parturition contributes significantly to the control of brucellosis,9 and, in addition, cows that abort should be isolated immediately and tested serologically soon after they aborted. If negative, they should be retested two to three weeks later. Animals testing positive should be culled. Foetuses, placentas and discharges must be disposed of, preferably by incineration.
It is important to maintain regular blood testing in infected herds.56 Serological tests at intervals of two to three months and bacteriological examination of aborted foetuses, placentas, lochia and milk are often of great benefit. Milk from negative animals should be tested regularly and at short intervals (monthly) by the MRT. All bulls from infected herds should be viewed with suspicion and the diagnosis should be based on clinical examination (including palpation of the seminal vesicles and ampullae), serological and allergenic tests, and on bacteriological examination of semen.160, 170
It is important to prevent, or, in outbreaks, at least limit the spread of the disease. Depending on circumstances on a farm, the availability of spare camps can be advantageous, particularly where it is deemed necessary to split the herd into two or more groups. The two-herd system, based on the segregation of weaner heifers and adults, can be highly effective in extensive farming operations. It is often not possible to cull all infected animals at the outset of an eradication campaign, but it is vitally important to separate them from the ‘negative’ herd. The creation of another ‘suspect’ group of animals, and the effective separation of cows from four months’ gestation onwards, may be beneficial. Replacement stock should be purchased from herds certified free of brucellosis.56
Whatever strategy is to be adopted, the owner must be fully aware of the implications of the proposed control programme. The cooperation of neighbouring farmers may be of assistance in this respect. In dealing with an infected herd, the need for decisive action is important. The ‘negative’ herd must be maintained as such and any contact with infected animals avoided. All bacteriologically or serologically positive animals must be permanently identified. Undoubtedly an element of ‘over-kill’ is built into a somewhat uncompromising and apparently ruthless approach in these programmes, but the long-term benefits will far outweigh this short-term disadvantage. The economic implications of a test-and-slaughter eradication policy must also be taken into account.9
If brucellosis is present in wildlife (see Brucella infections in terrestrial wildlife), the concern of the livestock owner is to prevent the reintroduction of the infection into his or her livestock, particularly in regions or states that are ‘officially brucellosis free’ because of the implications for pre-movement testing for domestic animals.24, 78 Decision-making to determine the importance of brucellosis control relative to other public and animal health concerns also needs to be addressed.129 in a “One Health” perspective.79
References
- AGUIRRE, N. P., VANZINI, V. R., TORIONI DE ECHAIDE, S., VALENTINI, B. S., DE LUCCA, G., AUFRANC, C., CANAL, A., VIGLIOCCO, A. & NIELSEN, K., 2002. Antibody dynamics in holstein friesian heifers vaccinated with Brucella abortusstrain 19, using seven serological tests. Journal of Immunoassay and Immunochemistry, 23, 471-478.
- ALEXANDER, B., SCHNURRENBERGER, P. R. & BROWN, R. R., 1981. Numbers of Brucella abortusin the placenta, umbilicus and foetal fluid of two naturally infected cows. Veterinary Record, 108, 500.
- ALTON, G. G., CORNER, L. A. & PLACKETT, P., 1980. Vaccination of pregnant cows with low doses of Brucella abortusstrain 19 vaccine. Australian Veterinary Journal, 56, 369-372.
- ALTON, G. G., JONES, L. M., ANGUS, R. D. & VERGER, J. M., 1988. Techniques for the Brucellosis Laboratory. 1st edn, Paris: Institut National de la Recherche Agronomique.
- ALTON, G. G., JONES, L. M., ANGUS, R. D. & VERGER, J. M., 1988. Techniques for the Brucellosis Laboratory. 1st edn. Paris: Institut National de la Recherche Agronomique.
- ALTON, G. G., JONES, L. M., ANGUS, R. D. & VERGER, J. M., 1988. Techniques for the Brucellosis Laboratory. lst edn. Paris: Institut National de la Recherche Agronomique.
- ANON., 1957. Obituary notice: L.E. Bevan, Veterinary Record,69, 421-422.
- ANON., 1986. Joint FAO/WHO Expert Committee on Brucellosis. Sixth Report. Geneva: World Health Organization.
- ANON., 1986. Joint FAO/WHO Expert Committee on Brucellosis. World Health Organization Technical Report Series. WHO, Geneva, 740.
- ANON., 1987. Directorate of Veterinary Services, South West Africa. Annual Report.
- ANON., 1988/89. Director of Animal Health, Directorate of Veterinary Services, Private Bag X138, Pretoria, South Africa. Annual Report.
- ANON., 1989/90. State Veterinarian, Skukuza. P.O. Box 12, Skukuza, South Africa. Annual Report.
- ANON., 1990. Brucellosis in Cattle (Contagious Abortion).Director of Animal Health, Directorate of Veterinary Services, Private Bag x138, Pretoria, South Africa.
- ANON., 2018. Brucellosis (Brucella abortus, B. melitensis and B. suis) (Infection with B. abortus, B. melitensis and B. suis). Manual of Standards for Diagnostic Tests and Vaccines. 8th edn. Paris: Office International des Epizooties.
- ANON., 2019. Manual of Standards for Diagnostic Tests and Vaccines. 3rd edn. Paris: Office International des Epizooties. https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.04_BRUCELLOSIS.pdf.
- ARAYA, L. N., ELZER, P. H., ROWE, G. E., ENRIGHT, F. M. & WINTER, A. J., 1989. Temporal development of protective cell-mediated and humoral immunity in BALB/c mice infected with Brucella abortus. Journal of Immunology,143, 3330-3337.
- ARAYA, L. N. & WINTER, A. J., 1990. Comparative protection of mice against virulent and attenuated strains of Brucella abortusby passive transfer of immune T cells or serum. Infection and Immunity, 58, 254-256.
- BALDWIN, C. L. & PARENT, M., 2002. Fundamentals of host immune response against Brucella abortus:What the mouse model has revealed about control of infection. Veterinary Microbiology, 90, 367-382.
- BANAI, M., ADAMS, L. G., FREY, M., PUGH, R. & FICHT, T. A., 2002. The myth of Brucella L-forms and possible involvement of Brucellapenicillinbinding proteins (PBPs) in pathogenicity. Veterinary Microbiology, 90, 263-279.
- BARTHEL, R., FENG, J., PIEDRAHITA, J. A., MCMURRAY, D. N., TEMPLETON, J. W. & G., A. L., 2001. Stable transfection of the bovine NRAMP1 gene into murine RAW264.7 Cells: Effect on Brucella abortusSurvival. Infection and Immunity, 69, 3110-3119.
- BARTON, C. E. & LOMME, J. R., 1980. Reduced-dose whole herd vaccination against brucellosis: A review of recent experience. Journal of the American Veterinary Medical Association, 177, 1218-1220.
- BARUCH, J., SUANES, A., PIAGGIO, J. M. & GIL, A. D., 2020. Analytic Sensitivity of an ELISA Test on Pooled Sera Samples for Detection of Bovine Brucellosis in Eradication Stages in Uruguay. Frontiers in Veterinary Science, 7, 178.
- BECKETT, F. W. & MACDIARMID, S. C., 1985. The effect of reduced-dose Brucella abortusstrain 19 vaccination in accredited dairy herds. British Veterinary Journal, 141, 507-514.
- BENGIS, R. G., KOCK, R. A. & FISCHER, J., 2002. Infectious animal diseases: the wildlife/livestock interface. Revue Scientifique et Technique de l’Office International des Epizooties, 21, 53-65.
- BISHOP, G. C., 1984. A brucellosis serological survey of beef cattle slaughtered at Cato Ridge Abattoir. Journal of the South African Veterinary Association,55, 185-186.
- BISHOP, G. C., 1990. Allerton Veterinary Laboratory, Pietermaritzburg, South Africa. Unpublished data.
- BLOOD, D. C., RADOSTITS, O. M. & HENDERSON, J. A., 1983. Veterinary Medicine. 6th edn. London: Baillière-Tindall.
- BRAUDE, A. I., 1951. Studies in the pathogenesis of experimental brucellosis. II. The formation of the hepatic granuloma and its evolution. Journal of Infectious Diseases,89, 87-94.
- BRICKER, B. J., 2002. PCR as a diagnostic tool for brucellosis. Veterinary Microbiology,90, 435-446.
- BRICKER, B. J. & HALLING, S. M., 1995. Enhancement of the Brucella AMOS PCR assay for differentiation of Brucella abortus vaccine strains S19 and RB51. Journal of Clinical Microbiology, 33, 1640-1642.
- BRICKER, B. J. & M., H. S., 1994. Differentiation of Brucella abortus (biovars 1, 2, and 4), Brucella melitensis, Brucella ovis, and Brucella suis (biovar 1) by the polymerase chain reaction. Journal of Clinical Microbiology, 32, 2660-2666.
- BRINLEY MORGAN, W. J., 1977. The diagnosis of Brucella abortusinfection in Britain. In: CRAWFORD, R. P. & HIDALGO, R. J., (eds). Bovine Brucellosis. An International Symposium. College Station, London: Texas A&M University Press.
- BUCK, J. M., 1930. Studies of vaccination during calfhood to prevent bovine infectious abortion. Journal of Agricultural Research, 41, 667-689.
- BYNDLOSS, M. X., TSAI, A. Y., WALKER, G. T., MILLER, C. N., YOUNG, B. M., ENGLISH, B. C., SEYFFERT, N., KERRINNES, T., DE JONG, M. F., ATLURI, V. L., WINTER, M. G., CELLI, J. & TSOLIS, R. M., 2019. Brucella abortus Infection of Placental Trophoblasts Triggers Endoplasmic Reticulum Stress-Mediated Cell Death and Fetal Loss via Type IV Secretion System-Dependent Activation of CHOP. mBio, 10, e01538-19.
- CAMPO, L. R., TAMAYO, R. & CAMPO, C. H., 1987. Embryo transfer from brucellosis-positive donors: A field trial. Theriogenology, 27, 221.
- CARGILL, C., LEE, K. & CLARKE, I., 1985. Use of an enzyme-linked immunoabsorbent assay in a bovine brucellosis eradication programme. Australian Veterinary Journal, 62, 49-52.
- CARON, E., GROSS, A., LIAUTARD, J. P. & DORNAND, J., 1996. Brucella species release a specific, protease-sensitive, inhibitor of TNF-alpha expression, active on human macrophage-like cells.Journal of Immunology, 156, 2885-2893.
- CATLIN, J. E. & SHEEHAN, E. J., 1986. Transmission of bovine brucellosisfrom dam to offspring. Journal of the American Veterinary Medical Association,188, 867-869.
- CELLI, J., 2015. The changing nature of the Brucella-containing vacuole. Cellular Microbiology, 17, 951-958.
- CELLI, J., 2019. The Intracellular Life Cycle of Brucella spp. Microbiology Spectrum, 7, 10.1128/microbiolspec.BAI-0006-2019.
- CHEVILLE, N., OLSEN, S., A., J., STEVENS, M., PALMER, M. & FLORANCE, A., 1996. Effects of age at vaccination on efficacy of Brucella abortusRB51 to protect cattle against brucellosis. American Journal of Veterinary Research, 57, 1153-1156.
- CLOECKAERT, A., JACQUES, I., BOSSERAY, N., LIMET, J. N., BOWDEN, R., DUBRAY, G. & PLOMMET, M., 1991. Protection conferred on mice by monoclonal antibodies directed against outer-membrane-protein antigens of Brucella. Journal of Medical Microbiology,34, 175-180.
- CLOECKAERT, A., JACQUES, I., DE WERGIFOSSE, P., DUBRAY, G. & LIMET, J. N., 1992. Protection against Brucella melitensisor Brucella abortus in mice with immunoglobulin G (IgG), IgA, and IgM monoclonal antibodies specific for a common epitope shared by the Brucella A and M smooth lipopolysaccharides. Infection and Immunity, 60, 312-315.
- CLOECKAERT, A., VERGER, J. M., GRAYON, M. & GREPINET, O., 1995. Restriction site polymorphism of the genes encoding the major 25 kDa and 36 kDa outer-membrane proteins of Brucella. Microbiology, 141, 2111-2121.
- CLOECKAERT, A., ZYGMUNT, M. S. & GUILLOTEAU, L. A., 2002. Brucella abortusvaccine strain RB51 produces low levels of M-like O-antigen. Vaccine, 20, 1820-1822.
- CONDY, J. B. & VICKERS, D. B., 1972. Brucellosis in Rhodesian wildlife. Journal of the South African Veterinary Medical Association, 43, 175-179.
- CONDY, L. B. & VICKERS, D. B., 1976. Brucellosis in buffalo (Syncerus caffer) in Wankie National Park. Rhodesian Veterinary Journal, 7, 58-60.
- CONSTANT, S. L. & BOTTOMLY, K., 1997. Induction of Th1 and Th2 CD4+ T cell responses: the alternative approaches. Annual Review of Immunology,15, 297-322.
- CORBEL, M. J., 1985. Recent advances in the study of Brucellaantigens and their serological cross-reactions. Veterinary Bulletin, 55, 927-942.
- CORBEL, M. J. & BRINLEY MORGAN, W. J., 1984. Genus Brucella Meyer and Shaw 1920.In: KRIEG, N. R. & HOLT, J. G., (eds). Bergey’s Manual of Systematic Bacteriology. Baltimore, London: Williams & Wilkins.
- CORBEL, M. J., STUART, F. A., BREWER, R. A., JEFFREY, M. & BRADLEY, R., 1989. Arthropathy associated with Brucella abortusstrain 19 vaccination in cattle. I. Examination of field cases. British Veterinary Journal, 145, 337-346.
- CORNER, L. A. & ALTON, G. G., 1981. Persistence of Brucella abortusstrain 19 infection in adult cattle vaccinated with reduced doses. Research in Veterinary Science, 31, 342-344.
- CORNER, L. A., ALTON, G. G. & IYER, H., 1987. Distribution of Brucella abortusin infected cattle. Australian Veterinary Journal, 64, 241-244.
- CRAWFORD, R. P., ADAMS, L. G., FICHT, T. A. & WILLIAMS, J. D., 1991. Effects of stage of gestation and breed on bovine responses to vaccination withBrucella abortus strain 19. American Journal of Veterinary Research, 199, 887-891.
- CRAWFORD, R. P., HUBER, J. D. & ADAMS, L. G., 1990. Epidemiology and surveillance.In: NIELSEN, K. & DUNCAN, B., (eds.). Animal Brucellosis. Orlando: CRC Press, 131-151.
- CRAWFORD, R. P., SANDERS, R. B. & ADAMS, L. G., 1988. Early removal of cows with brucellosis and the effect in strain 19 vaccinated cattle herds.Preventive Veterinary Medicine, 5, 191-195.
- CRAWFORD, R. P., WILLIAMS, J. D., D., H. J. & CHILDERS, A. B., 1979. Biotypes of Brucella abortusand their value in epidemiologic studies of infected cattle herds. Journal of the American Veterinary Medical Association, 175, 1274-1277.
- CUNNINGHAM, B., 1977. A difficult disease called brucellosis. In: CRAWFORD, R. P. & HIDALGO, R. J., (eds.). Bovine Brucellosis. An International Symposium.College Station, London: Texas A&M University Press.
- DA COSTA, M., GUILLOU, J. P., GARIN-BASTUJI, B., THIEBAUD, M. & DUBRAY, G., 1996. Specificity of six gene sequences for the detection of the genus Brucella by DNA amplification. Journal of Applied Bacteriology, 81, 267-275.
- DE VOS, V. & VAN NIEKERK, C. A. W. J., 1969. Brucellosis in the Kruger National Park. Journal of the South African Veterinary Medical Association, 40, 331-334.
- DETILLEUX, P. G., DEYOE, B. L. & CHEVILLE, N. F., 1990. Entry and intracellular localization ofBrucella spp. in Vero cells: Fluorescence and electron microscopy. Veterinary Pathology, 27, 317-328.
- DETILLEUX, P. G., DEYOE, B. L. & CHEVILLE, N. F., 1990. Penetration and intracellular growth of Brucella abortusin nonphagocytic cells in vitro. Infection and Immunity, 58, 2320-2328.
- DOLAN, L. A., 1980. Latent carriers of brucellosis. Veterinary Record, 106, 241-243.
- DOMENECH, J., LUCET, P., VALLAT, B., STEWART, C., BONNET, J. B. & BERTAUDIERE, L., 1980. La brucellose bovine en Afrique centrale. II. Etude clinique et épidémiologique: particularités régionales et problemes de l’élevage semi-intensif. Revue d’Elevage et de Médecine Vétérinaire des Pays Tropicaux,33, 277-284.
- DORNAND, J., GROSS, A., LAFONT, V., LIAUTARD, J., OLIARO, J. & LIAUTARD, J. P., 2002. The innate immune response against Brucella in humans. Veterinary Microbiology,90, 383-394.
- DUCROTOY, M. J., MUÑOZ, P. M., CONDE-ÁLVAREZ, R., BLASCO, J. M. & MORIYÓN, I., 2018. A systematic review of current immunological tests for the diagnosis of cattle brucellosis. Preventive Veterinary Medicine, 151, 57-72.
- DUFFIELD, B. J., STREETEN, T. A. & SPINKS, G. A., 1984. Isolation ofBrucella abortus from supramammary lymph nodes of cattle from infected herds vaccinated with low dose strain 19. Australian Veterinary Journal, 61, 411-412.
- ENRIGHT, F. M., 1990. The pathogenesis and pathobiology of Brucella infection in domestic animals. In: NIELSEN, K. & DUNCAN, B., (eds.). Animal brucellosis. Orlando: CRC Press, 301-320.
- EWALT, D. R., PAYEUR, J. P., RHYAN, J. C. & GEER, P. L., 1997. Brucella suisbiovar 1 in naturally infected cattle: a bacteriological, serological and histological study. Journal of Veterinary Diagnostic Investigation, 9, 417-420.
- FARRELL, I. D., 1974. The development of a new selective medium for the isolation of Brucella abortusfrom contaminated sources. Research in Veterinary Science, 16, 280-286.
- FAUL, A. & BOSMAN, P. P., 1987. South Africa. Bovine brucellosis and brucellosis of small ruminants. Technical Series, Office International des Epizooties, 6, 60-64.
- FENSTERBANK, R., 1978. Congenital brucellosis in cattle associated with localization in a hygroma. Veterinary Record,103, 283-284.
- FOSTER, J. T., BECKSTROM-STERNBERG, S. M., PEARSON, T., BECKSTROM-STERNBERG, J. S., CHAIN, P. S., ROBERTO, F. F., HNATH, J., BRETTIN, T. & KEIM, P., 2009. Whole-genome-based phylogeny and divergence of the genus Brucella. Journal of Bacteriology, 191, 2864-2870.
- FRETIN, D., MORI, M., CZAPLICKI, G., QUINET, C., MAQUET, B., GODFROID, J. & SAEGERMAN, C., 2013. Unexpected Brucella suis biovar 2 Infection in a dairy cow, Belgium. Emerging Infectious Diseases, 19(12), 2053-2054.
- GARDNER, I. A., HEITALA, S. & BOYCE, W. M., 1996. Validity of using serological tests for diagnosis of diseases in wild animals. Revue Scientifique et Technique de l’Office International des Epizooties,15, 323-335.
- GARIN, B., TRAP, D. & GAUMONT, R., 1985. Assessment of the EDTA seroagglutination test for the diagnosis of bovine brucellosis. Veterinary Record,117, 444-445.
- GODFROID, F., CLAVAREAU, C., TAMINIAU, B., DANESE, I., TIBOR, A., WEYNANTS, V., MERTENS, P., DE BOLLE, X., GODFROID, J. & LETESSON, J. J., 1998. Identification of the perosamine synthetase gene of Brucella melitensis16M and the involvement of the lipopolysaccharide O-side chain in Brucella survival in mice and in macrophages. Infection and Immunity, 66, 5485-5493.
- GODFROID, J., 2002. Brucellosis in wildlife. Revue Scientifique et Technique de l’Office International des Epizooties,21, 277-286.
- GODFROID, J., AL DAHOUK, S., PAPPAS, G., ROTH, F., MATOPE, G., MUMA, J., MARCOTTY, T., PFEIFFER, D. & SKJERVE, E., 2013. A "One Health" surveillance and control of brucellosis in developing countries: moving away from improvisation. Comparative Immunology, Microbiology and Infectious Diseases, 36(3), 241-248.
- GODFROID, J., GARIN-BASTUJI, B., SAEGERMAN, C. & BLASCO, J. M., 2013. Brucellosis in terrestrial wildlife. Revue Scientifique et Technique (Office International des Epizooties), 32, 27-42.
- GODFROID, J. & KASBOHRER, A., 2002. Brucellosis in the European Union and Norway at the turn of the twenty-first century. Veterinary Microbiology,90, 135-145.
- GODFROID, J., SAEGERMAN, C., WELLEMANS, V., WALRAVENS, K., LETESSON, J. J., TIBOR, A., MCMILLAN, A., SPENCER, S., SANNA, M., BAKKER, D., POUILLOT, R. & GARIN-BASTUJI, B., 2002. How to substantiate eradication of bovine brucellosis when aspecific serological reactions occur in the course of brucellosis testing. Veterinary Microbiology,90, 461-477.
- GODOY, A. M., PERES, J. N., BARG, L., COSTA, J. O. & DE GODOY, A. M., 1985. Ixodidae as a reservoir of animal brucellosis. Arquivo Brasileiro de Medicina Veterinaria e Zootecnia, 37, 369-375.
- GORVEL, J. P. & MORENO, E., 2002. Brucellaintracellular life: from invasion to intracellular replication. Veterinary Microbiology, 90, 281-297.
- GRADWELL, D. V., SCHUTTE, A. P., VAN NIEKERK, C. A. W. J. & ROUX, D. J., 1977. The isolation of Brucella abortusbiotype I from African buffalo in the Kruger National Park. Journal of the South African Veterinary Association, 48, 41-43.
- GROSS, A., TERRAZA, A., OUAHRANI-BETTACHE, S., LIAUTARD, J. P. & DORNAND, J., 2000. In vitro Brucella suisinfection prevents the programmed cell death of human monocytic cells. Infection and Immunity, 68, 342-351.
- HARMON, B. G., ADAMS, L. G., TEMPLETON, J. W. & SMITH, R., 1989. Macrophage function in mammary glands of Brucella abortus-infected cows and cows that resisted infection after inoculation of Brucella abortus.American Journal of Veterinary Research, 50, 459-465.
- HENNING, M. W., 1956. Animal Diseases in South Africa. 3rd edn. Pretoria, South Africa: Central News Agency Ltd.
- HERR, S., 1989. Causes of foetal death in cattle. Zimbabwe Veterinary Journal,20, 167-168.
- HERR, S. & MARSHALL, C., 1981. Brucellosis in free-living African buffalo (Syncerus caffer): A serological survey. Onderstepoort Journal of Veterinary Research, 48, 133-134.
- HERR, S. & ROUX, D., 1981. The efficacy of bacteriological procedures for the isolation of Brucella abortus from abattoir material. Onderstepoort Journal of Veterinary Research, 48, 7-12.
- HERR, S. & TE BRUGGE, L. A., 1984. Experience with reduced dose Brucella abortusstrain 19 vaccination of adult cattle in infected herds in South Africa. Proceedings of the Thirteenth World Congress on Diseases of Cattle, 1, 359-365.
- HERR, S. & TE BRUGGE, L. A., 1985. Profiles of serological reactions following adult cow inoculation with standard dose Brucella abortusstrain 19 vaccine. Journal of the South African Veterinary Association, 56, 93-96.
- HERR, S., TE BRUGGE, L. A. & GUINEY, M. C. M., 1982. The value of the microtitre serum agglutination test as a second screening test in bovine brucellosis. Onderstepoort Journal of Veterinary Research, 49, 23-28.
- HERR, S., WILLIAMSON, C. C., PRIGGE, R. E. & VAN WYK, A., 1986. The relationship between the microtitration serum agglutination test and complement fixation tests in bovine brucellosis serology. Onderstepoort Journal of Veterinary Research,53, 199-200.
- HOFFMAN, T., ROCK, K., MUGIZI, D. R., MURADRASOLI, S., LINDAHL-RAJALA, E., ERUME, J., MAGNUSSON, U., LUNDKVIST, Å. & BOQVIST, S., 2016. Molecular detection and characterization of Brucella species in raw informally marketed milk from Uganda. Infection Ecology and Epidemiology, 6, 32442.
- JAMIL, T., MELZER, F., SAQIB, M., SHAHZAD, A., KHAN, K. K., HAMMAD, H. M., RASHID, I., TAHIR, U., KHAN, I., HALEEM, T. M., ULLAH, S., MOHSIN, M., MANSOOR, M. K., SCHWARZ, S. & NEUBAUER, H., 2020. Serological and Molecular Detection of Bovine Brucellosis at Institutional Livestock Farms in Punjab, Pakistan. International Journal of Environmental Research and Public Health, 17(4), 1412.
- JONKER, J., 1988. Directorate of Animal Health, Eastern Cape Province and Karoo, Private Bag x12999, Centrahill. Personal communication.
- JUBB, K. V. F., KENNEDY, P. C. & PALMER, N., 1993. Pathology of Domestic Animals. 2nd edn. San Diego: Academic Press.
- JUBB, K. V. F., KENNEDY, P. C. & PALMER, N., 1993. Pathology of Domestic Animals. 4th edn. San Diego, California: Academic Press.
- KE, Y., WANG, Y., LI, W. & CHEN, Z., 2015. Type IV secretion system of Brucella spp. and its effectors. Frontiers in Cellular and Infection Microbiology, 5, 72.
- KEEP, M. E. & BISHOP, G. C., 1985. A brucellosis survey in Natal wildlife. Lammergeyer,35, 47-48.
- KEPPIE, J., WILLIAMS, A. E., WITT, K. & SMITH, H., 1965. The role of erythritol in the tissue localization of the brucellae. British Journal of Experimental Pathology, 46, 104-108.
- KOHLER, S., FOULONGNE, V., OUAHRANI-BETTACHE, S., BOURG, G., TEYSSIER, J., RAMUZ, M. & LIAUTARD, J. P., 2002. The analysis of the intramacrophagic virulome ofBrucella suisdeciphers the environment encountered by the pathogen inside the macrophage host cell. Proceedings of the National Academy of Sciences of the USA, 99, 15711-15716.
- KOHLER, S., LAYSSAC, M., NAROENI, A., GENTSCHEV, I., RITTIG, M. & LIAUTARD, J. P., 2001. Secretion of listeriolysin by Brucella suisinhibits its intramacrophagic replication. Infection and Immunity, 69, 2753-2756.
- KOHLER, S., PORTE, F., JUBIER-MAURIN, V., OUAHRANI-BETTACHE, S., TEYSSIER, J. & LIAUTARD, J. P., 2002. The intramacrophagic environment of Brucella suisand bacterial response. Veterinary Microbiology, 90, 299-309.
- KOLO, F. B., ADESIYUN, A. A., FASINA, F. O., KATSANDE, C. T., DOGONYARO, B. B., POTTS, A., MATLE, I., GELAW, A. K. & VAN HEERDEN, H., 2019. Seroprevalence and characterization of Brucella species in cattle slaughtered at Gauteng abattoirs, South Africa. Veterinary Medicine and Science, 5, 545-555.
- LA PRAIK, R. D., BROWN, D. D., MANN, H. & BRAND, T., 1975. Brucellosis: A study of five calves from reactor dams. Veterinary Record,97, 52-54.
- LA PRAIK, R. D. & MOFFAT, R., 1982. Latent bovine brucellosis. Veterinary Record, 111, 578-579.
- LE FLECHE, P., JACQUES, I., GRAYON, M., AL DAHOUK, S., BOUCHON, P., DENOEUD, F., NÖCKLER, K., NEUBAUER, H., GUILLOTEAU, L. A. & VERGNAUD, G., 2006. Evaluation and selection of tandem repeat loci for a Brucella MLVA typing assay. BMC Microbiology, 6, 9.
- LEDWABA, M. B., GOMO, C., LEKOTA, K. E., LE FLÈCHE, P., HASSIM, A., VERGNAUD, G. & VAN HEERDEN, H., 2019. Molecular characterization of Brucella species from Zimbabwe. PLoS Neglected Tropical Diseases, 13, e0007311.
- LETESSON, J. J., BARBIER, T., ZÚÑIGA-RIPA, A., GODFROID, J., DE BOLLE, X. & MORIYÓN, I., 2017. Brucella Genital Tropism: What's on the Menu. Frontiers in Microbiology, 8, 506.
- LETESSON, J. J., TIBOR, A., VAN EYNDE, G., WANSARD, V., WEYNANTS, V., DENOEL, P. & SAMAN, E., 1997. Humoral immune responses of Brucella-infected cattle, sheep, and goats to eight purified recombinant Brucellaproteins in an indirect enzyme-linked immunosorbent assay. Clinical and Diagnostic Laboratory Immunology, 4, 556-564.
- LEVIER, K., PHILLIPS, R. W., GRIPPE, V. K., ROOP, R. M. & WALKER, G. C., 2000. Similar requirements of a plant symbiont and a mammalian pathogen for prolonged intracellular survival. Science, 287, 2492-2493.
- LISTA, F., REUBSAET, F. A., DE SANTIS, R., PARCHEN, R. R., DE JONG, A. L., KIEBOOM, J., VAN DER LAAKEN, A. L., VOSKAMP-VISSER, I. A., FILLO, S., JANSEN, H. J., VAN DER PLAS, J. & PAAUW, A., 2011. Reliable identification at the species level of Brucella isolates with MALDI-TOF-MS. BMC Microbiology, 11, 267.
- LÓPEZ-GOÑI, I., GARCÍA-YOLDI, D., MARÍN, C. M., DE MIGUEL, M. J., MUÑOZ, P. M., BLASCO, J. M., JACQUES, I., GRAYON, M., CLOECKAERT, A., FERREIRA, A. C., CARDOSO, R., CORRÊA DE SÁ, M. I., WALRAVENS, K., ALBERT, D. & GARIN-BASTUJI, B., 2008. Evaluation of a multiplex PCR assay (Bruce-ladder) for molecular typing of all Brucella species, including the vaccine strains. Journal of Clinical Microbiology, 46, 3484-3487
- LÓPEZ–GOÑI, I., GARCÍA–YOLDI, D., MARÍN, C. M., DE MIGUEL, M. J., BARQUERO-CALVO, E., GUZMÁN-VERRI, C., ALBERT, D. & GARIN-BASTUJI, B., 2011. New Bruce-ladder multiplex PCR assay for the biovar typing of Brucella suis and the discrimination of Brucella suis and Brucella canis. Veterinary Microbiology, 154, 152-155.
- LORD, V. R., SCHURIG, G. G., CHERWONOGRODZKY, J. W., MARCANO, M. J. & MELENDEZ, G. E., 1998. Field study of vaccination of cattle with Brucella abortusstrains RB51 and 19 under high and low disease prevalence. American Journal of Veterinary Research, 59, 1016-1020.
- LUCERO, N. E., AYALA, S. M., ESCOBAR, G. I. & JACOB, N. R., 2008. Brucella isolated in humans and animals in Latin America from 1968 to 2006. Epidemiology and Infection, 136, 496-503.
- LUCHSINGER, D. W. & ANDERSON, R. K., 1979. Longitudinal studies of naturally acquired Brucella abortusinfection in sheep. American Journal of Veterinary Research, 40, 1307-1312.
- MACDIARMID, S. C. & HELLSTROM, J. S., 1987. An intradermal test for the diagnosis of brucellosis in extensively managed herds. Preventive Veterinary Medicine, 4, 361-369.
- MACMILLAN, A. P. & COCKREM, D. S., 1985. Reduction of non-specific reactions to the Brucella abortusserum agglutination test by the addition of EDTA. Research in Veterinary Science, 38, 288-291.
- MADSEN, M., 1989. The current state of brucellosis in Zimbabwe.Zimbabwe Veterinary Journal, 20, 133-148.
- MADZINA, W. M., 1987. Zimbabwe. Bovine brucellosis and brucellosis of small ruminants: diagnosis, control and vaccination. Technical Series, Office de l’International des Epizooties, 75017 Paris, France, 6, 80-82.
- MADZINGIRA, O., FASINA, F. O., KANDIWA, E., MUSILIKA-SHILONGO, A., CHITATE, F. & VAN HEERDEN, H., 2020. A retrospective sero-epidemiological survey of bovine brucellosis on commercial and communal farming systems in Namibia from 2004 to 2018. Tropical Animal Health and Production, 10.1007/s11250-020-02332-4.
- MANTHEI, C. A., DETRAY, D. E. & GOODE, E. R., 1950. Brucellainfection in bulls and the spread of brucellosis in cattle by artificial insemination. Proceedings of the 87th Annual Meeting of the American Veterinary Medical Association, 87, 177.
- MATHEW, C., STOKSTAD, M., JOHANSEN, T. B., KLEVAR, S., MDEGELA, R. H., MWAMENGELE, G., MICHEL, P., ESCOBAR, L., FRETIN, D. & GODFROID, J., 2015. First isolation, identification, phenotypic and genotypic characterization of Brucella abortus biovar 3 from dairy cattle in Tanzania. BMC Veterinary Research, 11, 156.
- MATOPE, G., BHEBHE, E., MUMA, J. B., SKJERVE, E. & DJØNNE, B., 2009. Characterization of some Brucella species from Zimbabwe by biochemical profiling and AMOS-PCR. BMC Research Notes, 2, 261.
- MCDERMOTT, J. J. & ARIMI, S. M., 2002. Brucellosis in sub-Saharan Africa: epidemiology, control and impact. Veterinary Microbiology, 90, 111-134.
- MCEWAN, A. D., 1937. Experiments on contagious abortion. II. Field immunization experiments with a vaccine prepared from strain 45. Veterinary Record, 49, 1585.
- MEADOR, V. P., DEYOE, B. L. & CHEVILLE, N. F., 1989. Pathogenesis of Brucella abortusinfection of the mammary gland and supramammary lymph node of the goat. Veterinary Pathology, 26, 357-368.
- MENSHAWY, A. M., PEREZ-SANCHO, M., GARCIA-SECO, T., HOSEIN, H. I., GARCÍA, N., MARTINEZ, I., SAYOUR, A. E., GOYACHE, J., AZZAM, R. A., DOMINGUEZ, L. & ALVAREZ, J., 2014. Assessment of genetic diversity of zoonotic Brucella spp. recovered from livestock in Egypt using multiple locus VNTR analysis. BioMed Research International, 353876.
- MOHAN, K., MAKAYA, P. V., MUVAVARIRWA, P., MATOPE, G., MAHEMBE, E. & PAWANDIWA, A., 1996. Brucellosis surveillance and control in Zimbabwe: bacteriological and serological investigation in dairy herds. Onderstepoort Journal of Veterinary Research, 48, 63, 47-51.
- MOSMANN, T. R. & SAD, S., 1996. The expanding universe of T-cell subsets: Th1, Th2 and more.Immunology Today, 17, 138-146.
- MUENDO, E. N., MBATHA, P. M., MACHARIA, J., ABDOEL, T. H., JANSZEN, P. V., PASTOOR, R. & SMITS, H. L., 2012. Infection of cattle in Kenya with Brucella abortus biovar 3 and Brucella melitensis biovar 1 genotypes. Tropical Animal Health and Production, 44(1), 17-20.
- MUMA, J. B., SYAKALIMA, M., MUNYEME, M., ZULU, V. C., SIMUUNZA, M. & KURATA, M., 2013. Bovine tuberculosis and brucellosis in traditionally managed livestock in selected districts of southern province of Zambia. Veterinary Medicine International, 730367.
- MURPHY, E. A., SATHIYASEELAN, J., PARENT, M. A., ZOU, B. & BALDWIN, C. L., 2001. Interferon-gamma is crucial for surviving a Brucella abortusinfection in both resistant C57BL/6 and susceptible BALB/c mice. Immunology, 103, 511-518.
- MUSALLAM, I., ABO-SHEHADA, M., HEGAZY, Y., HOLT, H. & GUITIAN, F., 2016. Systematic review of brucellosis in the Middle East: Disease frequency in ruminants and humans and risk factors for human infection. Epidemiology and Infection, 144(4), 671-685.
- MUSALLAM, I., NDOUR, A. P., YEMPABOU, D., NGONG, C. C., DZOUSSE, M. F., MOUICHE-MOULIOM, M. M., FEUSSOM, J., NTIRANDEKURA, J. B., NTAKIRUTIMANA, D., FANE, A., DEMBELE, E., DOUMBIA, A., AYIH-AKAKPO, A., PATO, P., PALI, M., TAPSOBA, A., COMPAORE, G. M., GAGARA, H., GARBA, A. I., CHENGAT PRAKASHBABU, B. & GUITIAN, J., 2019. Brucellosis in dairy herds: A public health concern in the milk supply chains of West and Central Africa. Acta Tropica, 197, 105042.
- NAIMAN, B. M., BLUMERMAN, S., ALT, D., BOLIN, C. A., BROWN, R., ZUERNER, R. & BALDWIN, C. L., 2002. Evaluation of type 1 immune response in naive and vaccinated animals following challenge withLeptospira borgpetersenii serovar hardjo: involvement of WC1(+) gammadelta and CD4 T cells. Infection and Immunity, 70, 6147-6157.
- NEGRÓN, M. E., KHAROD, G. A., BOWER, W. A. & WALKE, H., 2019. Notes from the Field: Human Brucella abortus RB51 Infections Caused by Consumption of Unpasteurized Domestic Dairy Products - United States, 2017-2019. MMWR, Morbidity and Mortality Weekly Report, 68, 185.
- NICOLETTI, P., 1980. The epidemiology of bovine brucellosis.Advances in Veterinary Science and Comparative Medicine, 24, 69-98.
- NICOLETTI, P., 1981. Prevalence and persistance of Brucella abortusstrain 19 infections and prevalence of other biotypes in vaccinated adult dairy cattle. Journal of the American Veterinary Medical Association, 178, 143-145.
- NICOLETTI, P., 1984. Vaccination of cattle with Brucella abortusstrain 19 administered by different routes and doses. Vaccine, 2, 133-135.
- NICOLETTI, P., 1990. Vaccination. In: NIELSEN, K. & DUNCAN, B., (eds.). Animal brucellosis. Orlando: CRC Press, 283-296.
- NICOLETTI, P., JONES, L. M. & BERMAN, D. T., 1978. Comparison of the subcutaneous and conjunctival route of vaccination with Brucella abortusstrain 19 vaccine in adult cattle. Journal of the American Veterinary Medical Association, 173, 1450-1456.
- NIELSEN, K., 1990. The serological response of cattle immunized with Yersinia enterocoliticaO:9 or O:16 to Yersinia and Brucella abortus antigens in enzyme immunoassays. Veterinary Immunology and Immunopathology, 24, 373-382.
- NIELSEN, K., 2002. Diagnosis of brucellosis by serology. Veterinary Microbiology,90, 447-459.
- NIELSEN, K. & DUNCAN, J. R., 1988. Antibody Isotype Response in Adult Cattle Vaccinated with Brucella abortusS19. Veterinary Immunology and Immunopathology, 19, 205-214.
- NIELSEN, K. & GALL, D., 2001. Fluorescence polarization assay for the diagnosis of brucellosis: A review. Journal of Immunoassay and Immunochemitry, 22, 183-201.
- NIELSEN, K., KELLY, L., GALL, D., NICOLETTI, P. & KELLY, W., 1995. Improved competitive enzyme immunoassay for the diagnosis of bovine brucellosis.Veterinary Immunology and Immunopathology, 46, 285-291.
- NIELSEN, K., SMITH, P., YU, W., NICOLETTI, P., JUNGERSEN, G., STACK, J. & GODFROID, J., 2006. Serological discrimination by indirect enzyme immunoassay between the antibody response to Brucella sp. and Yersinia enterocolitica O:9 in cattle and pigs. Veterinary Immunology and Immunopathology, 109, 69-78.
- OIE., 2020. https://www.oie.int/wahis_2/public/wahid.php/Diseaseinformation/Diseasetimelines
- OLIVEIRA, S. C., SOEURT, N. & SPLITTER, G., 2002. Molecular and cellular interactions between Brucella abortusantigens and host immune responses. Veterinary Microbiology, 90, 417-424.
- OTTONES, F., DORNAND, J., NAROENI, A., LIAUTARD, J. P. & FAVERO, J., 2000. V gamma 9V delta 2 T cells impair intracellular multiplication of Brucella suisin autologous monocytes through soluble factor release and contact-dependent cytotoxic effect. Journal of Immunology, 165, 7133-7139.
- PALMER, M. V., CHEVILL, E. N. F. & JENSEN, A. E., 1996. Experimental infection of pregnant cattle with the vaccine candidate Brucella abortusstrain RB51: pathologic, bacteriologic, and serologic findings. Veterinary Pathology, 33, 682-691.
- PALMER, M. V., OLSEN, S. & CHEVILLE, N. F., 1997. Safety and immunogenicity ofBrucella abortus RB51 vaccine in pregnant cattle. American Journal of Veterinary Research, 5, 472.
- PAWESKA, J. T., POTTS, A. D., HARRIS, H. J., SMITH, S. J., VILJOEN, G. J., DUNGU, B., BRETT, O. L., BUBB, M. & PROZESKY, L., 2002. Validation of an indirect enzyme-linked immunosorbent assay for the detection of antibody against Brucella abortusin cattle sera using an automated ELISA workstation. Onderstepoort Journal of Veterinary Research, 69, 61-77.
- PIZARRO-CERDA, J., MERESSE, S., PARTON, R. G., VAN DER GOOT, G., SOLA-LANDA, A., LOPEZ-GONI, I., MORENO, E. & GORVEL, J. P., 1998. Brucella abortustransits through the autophagic pathway and replicates in the endoplasmic reticulum of nonprofessional phagocytes. Infection and Immunity, 66, 5711-5724.
- PLANT, J. W., CLAXTON, P. D., JAKOVLJEVIC, M. B. & DE SARAM, W., 1976. Brucella abortusinfection in the bull. Australian Veterinary Journal, 52, 17-20.
- PORTE, F., LIAUTARD, J. P. & KOHLER, S., 1999. Early acidification of phagosomes containing Brucella suisis essential for intracellular survival in murine macrophages. Infection and Immunity, 67, 4041-4047.
- PRAUD, A., DURÁN-FERRER, M., FRETIN, D., JAY, M., O'CONNOR, M., STOURNARA, A., TITTARELLI, M., TRAVASSOS DIAS, I. & GARIN-BASTUJI, B., 2016. Evaluation of three competitive ELISAs and a fluorescence polarisation assay for the diagnosis of bovine brucellosis. Veterinary Journal, 216, 38-44.
- PROBERT, W. S., SCHRADER, K. N., KHUONG, N. Y., BYSTROM, S. L. & GRAVES, M. H., 2004. Real-time multiplex PCR assay for detection of Brucella spp., B. abortus, and B. melitensis. Journal of Clinical Microbiology, 42, 1290-1293.
- QURESHI, T., TEMPLETON, J. W. & ADAMS, L. G., 1996. Intracellular survival of Brucella abortus,Mycobacterium bovis BCG, Salmonella dublin, and Salmonella typhimurium in macrophages from cattle genetically resistant to Brucella abortus. Veterinary Immunology and Immunopathology, 50, 55-65.
- RAN, X., CHENG, J., WANG, M., CHEN, X., WANG, H., GE, Y., NI, H., ZHANG, X. X. & WEN, X., 2019. Brucellosis seroprevalence in dairy cattle in China during 2008-2018: A systematic review and meta-analysis. Acta Tropica, 189, 117-123.
- RANKIN, J. E. F., 1965. Brucella abortusin bulls: A study of twelve naturally infected cases. Veterinary Record, 77, 132-135.
- RHODES, S. G., HEWINSON, R. G. & VORDERMEIER, H. M., 2001. Antigen recognition and immunomodulation by gamma delta T cells in bovine tuberculosis. Journal of Immunology, 166, 5604-5610.
- ROOP, R. M., ROBERTSON, G. T., FERGUSON, G. P., MILFORD, L. E., WINKLER, M. E. & WALKER, G. C., 2002. Seeking a niche: Putative contributions of the hfq and bacA gene products to the successful adaptation of the brucellae to their intracellular home. Veterinary Microbiology,90, 349-363.
- RYLATT, D. B., WYATT, D. M. & BUNDESEN, P. G., 1985. A competitive enzyme immunoassay for the detection of bovine antibodies to Brucella abortus using monoclonal antibodies. Veterinary Immunology and Immunopathology, 8, 261-271.
- SAEGERMAN, C., VO, T. K., DE WAELE, L., GILSON, D., BASTIN, A., DUBRAY, G., FLANAGAN, P., LIMET, J. N., LETESSON, J. J. & GODFROID, J., 1999. Diagnosis of bovine brucellosis by skin test: Conditions for the test and evaluation of its performance.Veterinary Record, 145, 214-218.
- SAMARTINO, L. E., FORT, M., GREGORET, R. & SCHURIG, G. G., 2000. Use of Brucella abortusvaccine strain RB51 in pregnant cows after calfhood vaccination with strain 19 in Argentina. Preventive Veterinary Medicine, 45, 193-199.
- SANGARI, F. J., AGUERO, J. & GARCIA-LOBO, J. M., 2000. The genes for erythritol catabolism are organized as an inducible operon in Brucella abortus. Microbiology, 146, 487-495.
- SANOGO, M., ABATIH, E., THYS, E., FRETIN, D., BERKVENS, D. & SAEGERMAN, C., 2013. Importance of identification and typing of Brucellae from West African cattle: a review. Veterinary Microbiology, 164, 202-211.
- SCHURIG, G. G., ROOP, R. M., BAGCHI, T., BOYLE, S., BUHRMAN, D. & SRIRANGANATHAN, N., 1991. Biological properties of RB51; a stable rough strain of Brucella abortus. Veterinary Microbiology,28, 171-188.
- SCHURIG, G. G., SRIRANGANATHAN, N. & CORBEL, M. J., 2002. Brucellosis vaccines: past, present and future. Veterinary Microbiology, 90, 479-496.
- SHOME, R., TRIVENI, K., SWATI, S., RANJITHA, S., KRITHIGA, N., SHOME, B. R., NAGALINGAM, M., RAHMAN, H. & BARBUDDHE, S. B., 2019. Spatial seroprevalence of bovine brucellosis in India—A large random sampling survey. Comparative Immunology, Microbiology and Infectious Diseases, 65, 124-127.
- SIMPSON, G., MARCOTTY, T., ROUILLE, E., CHILUNDO, A., LETTESON, J. J. & GODFROID, J., 2018. Immunological response to Brucella abortus strain 19 vaccination of cattle in a communal area in South Africa. Journal of the South African Veterinary Association, 89(0), e1-e7.
- SIMPSON, G., MARCOTTY, T., ROUILLE, E., MATEKWE, N., LETESSON, J. J. & GODFROID, J., 2018. Documenting the absence of brucellosis in cattle, goats and dogs in a "One Health" interface in the Mnisi community, Limpopo, South Africa. Tropical Animal Health and Production, 50, 903-906.
- SINGH, B. B., DHAND, N. K. & GILL, J. P., 2015. Economic losses occurring due to brucellosis in Indian livestock populations. Preventive Veterinary Medicine, 119(3-4), 211-215.
- STAMP, J. T., MCEWEN, A. D., WATT, J. A. A. & NISBET, D. I., 1950. Enzootic abortion in ewes. I. Transmission of the disease. Veterinary Record, 62, 251–254.
- STRINGFELLOW, D. A., WOLFE, D. F., MCGUIRE, J. A., LAUERMAN, L. A., GRAY, B. W. & SPARLING, P. H., 1986. Effects of embryo-freezing and thawing techniques on the survivability ofBrucella abortus. Theriogenology, 26, 553-560.
- SUTHERLAND, S. S., 1984. Evaluation of the enzyme-linked immunosorbent assay in the detection of cattle infected with Brucella abortus. Veterinary Microbiology,10, 23-32.
- SUTHERLAND, S. S., 1985. An enzyme-linked immunosorbent assay for the detection of Brucella abortusin cattle using monoclonal antibodies. Australian Veterinary Journal, 62, 264-268.
- TANNER, M., INLAMEIA, O., MICHEL, A., MAXLHUZA, G., PONDJA, A., FAFETINE, J., MACUCULE, B., ZACARIAS, M., MANGUELE, J., MOIANE, I. C., MARRANANGUMBE, A. S., MULANDANE, F., SCHÖNFELD, C., MOSER, I., VAN HELDEN, P. & MACHADO, A., 2015. Bovine Tuberculosis and Brucellosis in Cattle and African Buffalo in the Limpopo National Park, Mozambique. Transboundary and Emerging Diseases, 62, 632-638.
- TAYLOR, A. W. & MACDIARMID, A., 1949. The stability of the avirulent characters of Brucella abortusstrain 19 and 45/20 in lactating and pregnant cows. Veterinary Record, 61, 317-318.
- TEBUG, S. F., NJUNGA, G. R., CHAGUNDA, M. G., MAPEMBA, J. P., AWAH-NDUKUM, J. & WIEDEMANN, S., 2014. Risk, knowledge and preventive measures of smallholder dairy farmers in northern Malawi with regard to zoonotic brucellosis and bovine tuberculosis. The Onderstepoort Journal of Veterinary Research, 81, 10.4102/ojvr.v81i1.594.
- THOEN, C. O. & ENRIGHT, F., 1986. Brucella.In: GYLES, C. L. & THOEN, C. O., (eds.). Pathogenesis of Bacterial Infections in Animals. 1st edn. Ames, Iowa: Iowa State University Press.
- THOMSON, A., 1950. Experimental studies on the incubation period of infectious abortion in cattle. British Veterinary Journal, 106, 41-54.
- THYANGATHYANGA, G. A. A., 1987. Malawi. Bovine brucellosis and brucellosis of small ruminants: Diagnosis, control and vaccination. Technical Series, Office International des Epizooties, 6, 50-55.
- TRICHARD, C. J. V., HERR, S., BASTIANELLO, S. S. & ROUX, D., 1982. Unilateral orchitis in a bull caused byBrucella abortus Biotype 1. Journal of the South African Veterinary Association, 53, 60-62.
- VAN METRE, D. C., KENNEDY, G. A., OLSEN, S. C., HANSEN, G. R. & EWALT, D. R., 1999. Brucellosis induced by RB51 vaccine in a pregnant heifer. Journal of the American Veterinary Medical Association,215, 1491-1493.
- VANZINI, V. R., AGUIRRE, N., LUGARESI, C. I., DE ECHAIDE, S. T., DE CANAVESIO, V. G., GUGLIELMONE, A. A., MARCHESINO, M. D. & NIELSEN, K., 1998. Evaluation of an indirect ELISA for the diagnosis of bovine brucellosis in milk and serum samples in dairy cattle in Argentina. Preventive Veterinary Medicine, 36, 211-217.
- VANZINI, V. R., AGUIRRE, N. P., VALENTINI, B. S., TORIONI DE ECHAIDE, S., LUGARESI, C. I., MARCHESINO, M. D. & NIELSEN, K., 2001. Comparison of an indirect ELISA with the Brucella milk ring test for detection of antibodies to Brucella abortus in bulk milk samples. Veterinary Microbiology, 82, 55-60.
- VEMULAPALLI, R., MCQUISTON, J. R., SCHURIG, G. G., SRIRANGANATHAN, N., HALLING, S. M. & BOYLE, S. M., 1999. Identification of an IS711 element interrupting the wboA gene ofBrucella abortus vaccine strain RB51 and a PCR assay to distinguish strain RB51 from other Brucella species and strains. Clinical and Diagnostic Laboratory Immunology, 6, 760-764.
- VERGER, J. M., GARIN-BASTUJI, B., GRAYON, M. & MAHÉ, A. M., 1989. La brucellose bovine à Brucella melitensisen France. Annales de Recherche Vétérinaire, 20, 93-102.
- VERGNAUD, G., HAUCK, Y., CHRISTIANY, D., DAOUD, B., POURCEL, C., JACQUES, I., CLOECKAERT, A. & ZYGMUNT, M. S., 2018. Genotypic Expansion Within the Population Structure of Classical Brucella Species Revealed by MLVA16 Typing of 1404 Brucella Isolates From Different Animal and Geographic Origins, 1974-2006. Frontiers in Microbiology, 9, 1545.
- WEYNANTS, V., GILSON, D., CLOECKAERT, A., TIBOR, A., DENOEL, P. A., GODFROID, F., LIMET, J. N. & LETESSON, J. J., 1997. Characterization of smooth-lipopolysaccharide and O polysaccharides of Brucellaspecies by competition binding assays with monoclonal antibodies. Infection and Immunity, 65, 1939-1943.
- WEYNANTS, V., GODFROID, J., LIMBOURG, B., SAEGERMAN, C. & LETESSON, J. J., 1995. Specific bovine brucellosis diagnosis based on an in vitroantigeno-specific IFN-gamma production. Journal of Clinical Microbiology 33, 706-712.
- WEYNANTS, V., TIBOR., A., DENOEL, P. A., SAEGERMAN, C., GODFROID, J., THIANGE, P. & LETESSON, J. J., 1996. Infection of cattle with Yersinia enterocoliticaO:9 a cause of the false positive serological reactions in bovine brucellosis diagnostic tests. Veterinary Microbiology, 48, 101-112.
- WHATMORE, A. M. & GOPAUL, K. K., 2011. Recent advances in molecular approaches to Brucella diagnostics and epidemiology. In: Brucella: Molecular Microbiology and Genomics, LÓPEZ-GOÑI, I. & O’CALLAGHAN, D., (eds.). Caister Academic Press, Norfolk, UK, 57-88.
- WHATMORE, A. M. & GOPAUL, K. K., 2011. Recent advances in molecular approaches to Brucella diagnostics and epidemiology. In: LÓPEZ-GOÑI, I. & O’CALLAGHAN, D., (eds.). Caister Academic Press, Norfolk, UK. Brucella: Molecular Microbiology and Genomics, 57-88.
- WILESMITH, J. W., 1978. The persistence of Brucella abortusinfection in calves: A retrospective study of heavily infected herds. Veterinary Record, 103, 149-153.
- WILLIAMS, A. E., KEPPIE, J. & SMITH, H., 1962. The chemical basis of the virulence of Brucella abortus. III. Foetal erythritol, a cause of the localization ofBrucella abortus in pregnant cows. British Journal of Experimental Pathology, 43, 530-537.
- WRIGHT, A. E. & SMITH, F., 1897. On the application of the serum test to differential diagnosis of thyphoid and Malta fever. Lancet,1, 656-659.
- WYCKOFF, J. H., 2002. Bovine T lymphocyte responses to Brucella abortus. Veterinary Microbiology, 90, 395-415.
- WYNN-JONES, G., BAKER, J. R. & JOHNSON, P. M., 1980. A clinical and immunopathological study of Brucella abortusstrain 19-induced arthritis in cattle. Veterinary Record, 107, 5-9.
- XAVIER, M. N., PAIXAO, T. A., POESTER, F. P., LAGE, A. P. & SANTOS, R. L., 2009. Pathological, Immunohistochemical and Bacteriological Study of Tissues and Milk of Cows and Fetuses Experimentally Infected with Brucella abortus. Journal of Comparative Pathology, 140, 149-157.
- XAVIER, M. N., WINTER, M. G., SPEES, A. M., NGUYEN, K., ATLURI, V. L., SILVA, T. M., BÄUMLER, A. J., MÜLLER, W., SANTOS, R. L. & TSOLIS, R. M., 2013. CD4+ T cell-derived IL-10 promotes Brucella abortus persistence via modulation of macrophage function. PLoS Pathogens, 9(6), e1003454.
- XIN, X., 1986. Orally administrable brucellosis vaccine: Brucella suisstrain 2 vaccine. Vaccine, 4, 212-216.
- YAGER, J. A. & SCOTT, D. W., 1993. Skin and Appendages. In: JUBB, K. V. F., KENNEDY, P. C. & PALMER, N., (eds.). Pathology of Domestic Animals. 4th ed. New York, London: Academic Press.
- YOUNG, E. J., 1995. An overview of human brucellosis. Clinical Infectious Diseases, 21, 283-290.
- ZHAN, Y. & CHEERS, C., 1993. Endogenous gamma interferon mediates resistance to Brucella abortus infection. Infection and Immunity, 61, 4899-4901.
- ZHAN, Y. & CHEERS, C., 1995. Differential induction of macrophagederived cytokines by live and dead intracellular bacteria in vitro. Infection and Immunity,63, 720-723.
- ZHAN, Y. & CHEERS, C., 1995. Endogenous interleukin-12 is involved in resistance to Brucella abortusinfection. Infection and Immunity, 63, 1387-1390.
- ZHAN, Y., LIU, Z. & CHEERS, C., 1996. Tumor necrosis factor alpha and interleukin-12 contribute to resistance to the intracellular bacterium Brucella abortusby different mechanisms. Infection and Immunity, 64, 2782-2786.




