- Infectious Diseases of Livestock
- Part 3
- Proliferative enteropathies of pigs
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Proliferative enteropathies of pigs
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Proliferative enteropathies of pigs
Previous authors: G H K LAWSON AND M C WILLIAMS
Current authors:
M, JACOBSON, Professor, DVM, PhD, Dipl. ECPHM, Department of Clinical Sciences, SLU, Swedish University of Agricultural Sciences, Sweden
Introduction
Proliferative enteropathy (PE) is a pathological description of the underlying intestinal lesion that occurs in a number of clinical conditions in pigs. The clinical diseases are known by their pathological description. The term describes the lesion, which is a persistent proliferation of enterocytes, and which in its early stages is often not accompanied by marked evidence of intestinal inflammation. All cases of PE in pigs show the presence of curved intracellular bacteria located in the apical cytoplasm of proliferating cells. Some other porcine enteric diseases may show some short-lived, minor epithelial proliferation but an absence of intracellular bacteria.2 Other animal species may be affected by proliferative enteropathy with intracellular bacteria,22, 24, 27, 107, 149 while others may show proliferative enteropathy in the absence of intracellular bacteria.19 In all cases where enterocyte proliferation has been associated with intracellular bacteria, the evidence available to date suggests that the organism is Lawsonia intracellularis, or a closely related species.2
The disease in pigs differs clinically depending on the age of the affected pigs. Post-weaning animals fail to grow normally and show intermittent diarrhoea, and their condition is often described as porcine intestinal adenomatosis (PIA). Subclinical cases, manifesting as poor growth only, are commonly seen. Older animals are affected by a condition of intestinal blood loss referred to as proliferative haemorrhagic enteropathy (PHE). Changes superimposed on PIA can result in necrotic enteritis (NE) and subsequently in regional ileitis (RI).46, 129
Proliferative enteropathy was first described in North America in 1931,6 and now probably occurs wherever domestic pigs are reared.. In South Africa and probably elsewhere in Africa, the disease is still most commonly recognized as PHE, and occurs in large, intensive commercial units.29, 155, 158
Aetiology
The demonstration of bacteria within the cells of the lesions of PE131 initiated a search for this organism that proved confusing to all involved.74 The curved morphology of the intracellular bacteria indicated a possible relation with the genus Campylobacter and such bacteria could be recovered from the lesions.25, 33, 77, 143, 144 A variety of these organisms came to be associated with the disease, but none proved capable of reproducing the condition experimentally.9, 87, 122, 123 It was only when the intracellular bacteria proved antigenically dissimilar to the porcine intestinal campylobacters that evidence for the presence of another distinct agent began to emerge.78, 89, 90 The new organism, an obligate intracellular bacterium that could be co-cultivated with rat enterocytes,75 belonged to a new genus and, after a tortuous process, was named Lawsonia intracellularis.91 The intracellular organism had been described as Campylobacter-like organisms, intracellular organisms, or ileal symbiont intracellularis before the present name was approved.30
The bacteria are variably curved or sigmoid Gram-negative rods, 1.25 to 1.75 μm long and 0.25 to 0.43 μm wide. The bacterial envelope consists of a wavy trilaminar outer layer separated from the cytoplasmic membrane by an electron-lucent zone.30 Extracellularly, there is a single, unipolar flagellum.72 In the natural disease, bacteria are present as groups in the apical cytoplasm between the cell nucleus and the luminal border. They lie free in the cytoplasm and are not surrounded by a host cell membrane.128 The bacteria adopt a similar location in cultured cells.93
Bacteria have commonly been classified based on the sequences of the genes coding for the 16S rDNA.154 On the evidence of the base sequences of bacterial 16S rDNA and the GroEL amino acid sequences, L. intracellularis is grouped into the Desulfovibrionaceae family in the delta subdivision of the Proteobacteria, and is taxonomically isolated from other characterized bacteria.20, 30, 91 The closest relatives are Bilophila wadsworthia and Desulfovibrio desulfuricans.30, 137 To date, six whole-genome sequences of L. intracellularis have been published, namely the porcine isolate PHE/MN1-00, originating from a gilt with PHE (ATCC PTA-3457),35 the isolate N343 that came from a sow with PHE from Minnesota,135 three draft genome sequences of isolates from Japanese pigs with PIA,109 and one isolate from a horse with a 99.63 per cent similarity to the porcine isolates.104 Porcine strains from Europe, the USA68 and Australia16 appear similar in characteristics in culture and identification by PCR or immunohistochemistry. An 18 kb prophage-associated genomic island may be a feature for adaptation to the porcine host.147
The bacterium can be grown in a number of cell lines, notably the fast dividing Mc Coy cells (ATCC® CRL-1696 ™), rat small intestinal cell line IEC-18 (ATCC CRL 1589) and INT-407 Henle intestinal cells (ATCC CCL 6).16, 56, 75, 121 Infected pig tissue is homogenized and filtered to remove tissue debris and contaminating bacteria. The filtrate is then applied to partially grown monolayers of cells in the presence of vancomycin and neomycin in an atmosphere of nitrogen containing 6 to 8 per cent oxygen and 5 per cent carbon dioxide. The bacterium multiplies within the cells, which continue to divide. Heavily infected cells reach maximum numbers about seven to eight days after infection. Passage of the bacteria is achieved by lysis of infected cells and infection of fresh monolayers. Using these methods, L. intracellularis can be recovered and propagated from most recently dead pigs affected with PE.75 All attempts to grow the bacterium on conventional bacteriological media (including specialized media for enteric organisms) and embryonated eggs, have failed.30, 60
SDS-polyacrylamide gel electrophoresis and western blotting, using either polyclonal or monoclonal antisera, demonstrates a number of envelope antigens. Major bands have a molecular weight of 53 000 and 42 000 (53 and 42K) with a lesser band of 47K; two minor bands of 25 to 27K are recognized by a monoclonal antibody89, 90 and are probably located in the outer leaflet of the envelope.95 All strains of L. intracellularis appear to react with this latter reagent. Following publication of the whole genome sequence and by the use of proteomic techniques, several surface antigens have been identified: the surface antigen A Lsa A) is associated with cellular attachment and invasion;88 an ATP/ADP translocase;138 a putative haemolysin gene; ( LhlyA)65 a putative type 3 secretion system (T3SS);1 an autotransporter protein A ( LatA);151 a flagellar-associated protein (LI0570);157 and outer membrane proteins expressed during infection.151
Epidemiology
Herd infection is common (20 to 100 per cent) whatever the production system and management, although the prevalences vary depending on the diagnostic method applied.42, 47, 66, 116 The disease occurs throughout the world.6, 106, 114, 120
Disease in adult or mature animals (> 4 to 4.5 months of age) almost always manifests as PHE, with the pattern of infection resembling an acute onset infectious disease. The intestinal haemorrhage seem to be superimposed on the primary proliferative lesions.44, 53, 130 Herd morbidity is variable, but, where most animals are considered susceptible, may range from between 12 and 50 per cent of animals at risk; mortality in clinically affected animals is high (up to 50 per cent).38, 83 Porcine haemorrhagic enteropathy is a dramatic disease that is difficult to ignore. High health status farms have always been considered as likely to be affected by PHE, and this conclusion, although previously largely unquantified, has now been shown to have a statistical basis.140 In recently formed herds and animals in growth trials there are other circumstances that make the appearance of PHE likely,38, 140 asthey may lead to the separation of young growing stock from sources of infection, which thus develop into susceptible mature animals. The likely sources of infection are infected, non-clinical adults, infected faeces transferred by personnel, or other infected animal species.
It has been shown that during outbreaks the number of animals excreting the bacterium is greater than the number showing signs, and it is possible, therefore, that numerous subclinical infections occur in adult animals.14, 39 The presence of a similar disease in rabbits, deer, foxes, horses, ferrets and some avian species indicates that infection by bacteria closely related to L. intracellularis is, on occasion, present in many animal species.17, 22, 24, 27 Presence of bacterial DNA has been confirmed in rats and mice that often are co-habitants on pig farms.3, 28 However, infection in species other than pigs has not been incriminated yet as the origin of infection in pigs.
Some clinical outbreaks may be associated with suppression or elimination of infection by in-feed medication. Subsequent withdrawal of treatment and contact with other pigs may result in clinical disease.4, 64
The epidemiology of infection is better understood when it occurs in weaner pigs. In one study, the greatest number (22,9 per cent) of positive animals, as assessed by the PCR test on faeces, was found between 10 and 24 days after they were weaned. Growing animals yielded fewer positive samples (12.9 per cent), and adults were rarely positive (0 to 3.4 per cent).105 Similarly, in a longitudinal study 50 pigs were sampled every third week during eight months from weaning and onwards. Following weaning and transfer to a growing unit, all pigs seroconverted or shed the microbe, with peak shedding occurring three and six weeks after the introduction into the premises.46 Infection occurring in the growing or fattening period is associated with a variable appearance of clinical disease. Infection may be largely subclinical14, 34 or result in mortality of about 1 per cent of throughput,43, 80 generally associated with NE or RI. The observation that certain management systems appear to affect the occurrence of disease suggests that many of these infections are derived from persistent infection in the pig’s environment.46, 64, 140
Excretion of L. intracellularis, as assessed by the PCR test, is detected from seven days post- infection with peak excretion at two weeks after exposure.36 Shedding may be irregular but may persist for 10 to 12 weeks following experimental inoculation36, 139 and single animals may develop into chronic carriers of the infection.46
Pathogenesis
An understanding of the pathogenesis of PE has been derived not only from more recent experiments in pigs using cultured bacteria,98, 146, 151 but from much earlier experiments in both pigs84, 85, 125 and hamsters,49-51, 58, 125 using diseased tissue as inocula. Additional information has also been derived from observations of cell culture infection, which appears to possess many of the features of the natural disease.73, 75, 93, 97 Non-pathogenic isolates of L. intracellularis have been derived through multiple passages in cell cultures in vitro and comparisons have been made between pathogenic and non-pathogenic isolates at molecular level. The results demonstrated the repression of plasmid A in non-pathogenic isolates, suggesting a mechanism for gene silencing (switching off).145 However, the mechanisms behind the extensive cell proliferation that is a hallmark of the disease have not been fully elucidated.
Pigs exposed to PE tissue develop lesions of PIA, NE or PHE, and it is now generally accepted that all the clinical pathological manifestations are due to a single cause.84 Conventional pigs exposed to bacteria in pure culture develop PE, but gnotobiotic animals, either germ-free or those with a non-intestinal bacterial flora, are not susceptible to either disease or infection.92 Germ-free pigs seeded with intestinal bacteria are, however, susceptible to infection with L. intracellularis and lesions develop.98 Similarly, gnotobiotic piglets exposed to PE tissue filtrate containing residual intestinal bacteria also develop lesions.94 It thus appears that the presence of other intestinal bacteria non-specifically modifies susceptibility to infection.
The microbe seems to adapt to some extent to various host species . Hamsters are susceptible to infection by L. intracellularis derived from pigs and the sequence of events is very similar to hamsters exposed to hamster-derived bacteria.49 However, hamsters seem less susceptible to isolates derived from horses31, 51 Furthermore, rabbits do not develop typical lesions when challenged with a porcine isolate.136 Pigs develop a mild disease when exposed to a horse isolate and horses develop a mild disease when exposed to a pig isolate.148 However, rabbits and horses seem to be equally susceptible to horse and rabbit isolates, respectively.118
The infection is spread by the faecal-oral route. In cell culture, the extracellular unipolar flagellum gives rise to a darting motion of the bacterium that probably is important for colonization in the intestines.72 During infection, the bacterium synthesizes a protein involved in cell attachment and invasion.88 but cell entry seems dependent on host cell activity and actin polymerization.73 The early events of intestinal infection have been studied most thoroughly in hamsters. The bacteria are first seen in small intestinal crypt cells five days after infection, but they are not present on the cell surface. However, one study in pigs describes the presence of the bacterium within the epithelial cells at the tips of the villi, 12 hours after inoculation with a mucosal homogenate.10 In a pig ligated-intestinal-loop model, the bacterium was taken up by the enterocytes within 6 h post- infection.11 Ten days after exposure, hyperplastic changes in intestinal morphology associated with the presence of the intracellular bacteria are present. These consist of the extension of crypt-type epithelium onto the base of the villus walls. The spread of infected hyperplastic epithelium continues and reaches its maximum about 21 days post-infection.49 The development of cell hyperplasia closely follows the spread of infection throughout the epithelium.49
Organisms can be seen dividing within cells, but are not seen on the cell surface or passing between cells. These features indicate that colonization of the epithelium is mainly by the division of infected cells. In advanced or developed lesions, degenerate infected cells may be extruded from the epithelium and may then release their contained bacteria.100 Crypt cells free of infection and adjacent to lesions quickly revert to the normal pattern of differentiation, including goblet cell formation.100
Some evidence suggests that the immune response to L. intracellularis is not robust. Inflammatory cell responses are often minimal, even in developed lesions,44, 133 and many animals only show a marked and transient IgM response to the organism.76 More recent studies suggest that secretory IgA and cell- mediated immune responses play an important role in protective immunity.18, 37
Minor differences have been described in the immune cells of established lesions of PHE and PIA; CD8 and CD25 T cells are located in the lamina propria in both conditions, but IgM B cells are more numerous in PHE.96 Substantial accumulations of IgA are present in the epithelial cells at the site of the intracellular bacteria in all forms of PE.79, 96 Hypersensitivity-mediated responses have been suggested as the basis for the haemorrhagic blood loss,82 and investigations into field cases showed that PHE may be indicative of aggravated immune responses.44
Necrotic enteritis is characterized not only by necrosis but also by inflammation of the hyperplastic mucosa. It has been speculated that this occurs because the immature epithelium without its protective mucous coat is more susceptible to damage by other intestinal bacteria than is the mature epithelium.5, 129
Regional ileitis seems to be the most protracted form of the disease, judging by the often chronic, granulomatous inflammation that extends to the full depth of the submucosa. The cause of the marked hypertrophy of the intestinal muscle layers is unknown, but it may reflect the greater effort required to transfer ingesta through a severely damaged intestine.129
Clinical signs
Proliferative enteropathy presents a variable set of signs depending on the age of the affected animal and the complications superimposed on the primary lesion. The incubation period is one to three weeks.8, 139 Disease is most commonly seen three to four weeks post- weaning or in 10–12-week-old pigs141 although pigs may be affected at any time from 2–20 weeks of age.54, 156 In uncomplicated cases of the chronic form of the disease, PIA, the mortality is usually low (1–5 per cent) and mainly results from secondary infections.124, 156 In subclinical cases, poor growth is the only clinical sign.34, 124 More obvious cases of PIA are characterized by an initial period during which there may be protracted, mild diarrhoea that may be followed by wasting, anaemia, the appearance of a roughened hair coat and even emaciation.6, 57, 156 It is important to emphasize that even in well-managed enterprises, stockmen may be unaware of the presence of disease. Only close examination of the rate of growth will disclose the depressant effect of PE, and it is unusual for pigs to be weighed frequently enough to accurately indicate the problem. Most of the clinically affected animals recover within six weeks, after a remarkably sudden return of appetite, and thereafter progress normally.34, 124
Cases of NE occur sporadically in weaned pigs, mostly between the ages of three and six months.6, 7 Affected animals usually show only mild to moderate diarrhoea that lasts several weeks,6, 7, 21 and pieces of pseudomembrane reminiscent of those seen in chronic salmonellosis may be observed in the stools.62 The pigs gradually lose weight, become emaciated and eventually die.6, 7
Regional ileitis is a chronic manifestation of the disease and occurs only in weaned pigs. It is often asymptomatic and found incidentally in baconers at slaughter.23 Those that are clinically affected may show mild anorexia, weight loss, and either diarrhoea or constipation.23, 26, 86, 108 In single cases, the abdomen may be bloated due to a constriction of the intestine or perforation of the ileum may supervene, leading to peritonitis and sudden death.23, 26, 108 Terminal signs, if seen, are those of complete anorexia, vomiting, excessive thirst, dullness, and disinclination to move.
Porcine haemorrhagic enteropathy is the most serious manifestation of PE, as it usually occurs as explosive outbreaks in mature or adult pigs.38, 112, 113 On occasion, clinical signs have been recorded in pigs before or at weaning in association with disease in adults, but this is not a common occurrence.133 Occasionally, sudden death occurs without premonitory signs.83 Most animals that develop PHE become anorexic, show mucosal pallor, and have faeces that vary from merely dark or blood-stained to profuse dysentery.83, 111, 112, 120, 133 Some animals vomit, have laboured breathing, and heavily pregnant sows may abort.4, 83, 112 A few pigs are mildly pyrexic but most have normal to subnormal temperatures.83, 112, 120, 133 Animals that survive recover slowly over a period of a week.
Pathology
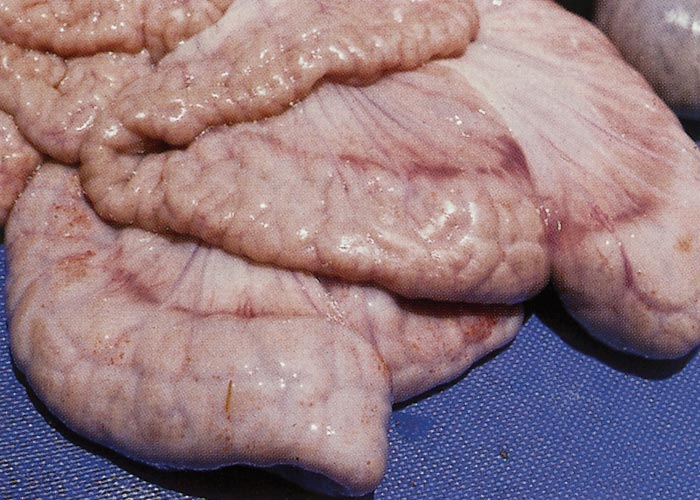
The carcasses of animals affected by uncomplicated PIA vary in condition. Severe lesions are often associated with wasted or emaciated carcasses;71 minor lesions, revealed only by microscopic examination, may have little effect on bodily condition.124, 139 The most characteristic lesion occurs in the mucosa of the terminal ileum,21, 71 and less often in the caecum and proximal colon.6, 71 In a few cases just the large intestine, including the distal colon, is affected.6, 52 Severely affected ilea are distended, and there is variable subserosal oedema.133 The affected mucosa is markedly thickened and thrown into interconnecting longitudinal and transverse folds up to 5 mm wide and 4 mm high (Figure 1).21, 71 This thickening is usually diffuse, but may have a nodular pattern;71 it is common to find nodular lesions in the more proximal ileum and diffuse lesions in the lower levels. Nodules are 2 to 15 mm in diameter, are raised above the adjacent mucosal surface and, in some cases, resemble polyps, as they are often wider at the surface than at the base.6, 21, 133 The mucosa is greyish-white, and small areas of superficial necrosis infrequently occur.71 Necrosis is probably indicative of a transitional phase between PIA and NE. The mesenteric lymph nodes that drain affected portions of the intestine are usually enlarged.6
Necrotic enteritis is characterized by intestinal lesions that show a similar distribution to those of PIA but, in addition, the surface epithelium or much of the entire mucosa is replaced by a diphtheritic pseudomembrane or necrotic tissue;6, 7, 127, 129 this altered mucosa is grey-yellow and deeply fissured. Oedema of the intestinal wall and of the mesentery may be observed in some instances.129 Pigs that died as a result of NE are invariably emaciated.6, 7
When RI is an incidental finding at slaughter, the carcass is usually well fleshed,26 but in fatal field cases the animal is often emaciated.62 Marked thickening and rigidity (‘hosepipe’ gut) of the wall of the terminal ileum is the characteristic gross finding,23, 26, 62, 108, 119, 129 the external diameter of the intestine being up to three times that of normal pigs.23 The increase in diameter is due to severe hypertrophy of the longitudinal and circular muscle layers of the tunica muscularis and, in most cases, concomitant thickening of the mucosa.23, 26, 62, 108, 119, 129 The serosa of the terminal ileum and associated mesentery may be both thickened and oedematous23, 62 or congested.23, 119 In some cases there is a patchy, fibrinous, granulomatous or fibrous serositis.23, 119 The mucosa may be whitish, greyish, yellowish or diffusely reddened,23 and occasionally there are ulcers covered with necrotic debris and pseudomembranes.23, 62, 119, 129 Peyer’s patches and the ileo-caecal lymph nodes may be hyperplastic,23, 26 and ileo-caecal lymphadenitis has also been described.23 If perforation of the ileum occurs, it usually takes place at the attachment of the meso-ileum23 and almost invariably results in diffuse fibrinous peritonitis.23, 26, 108
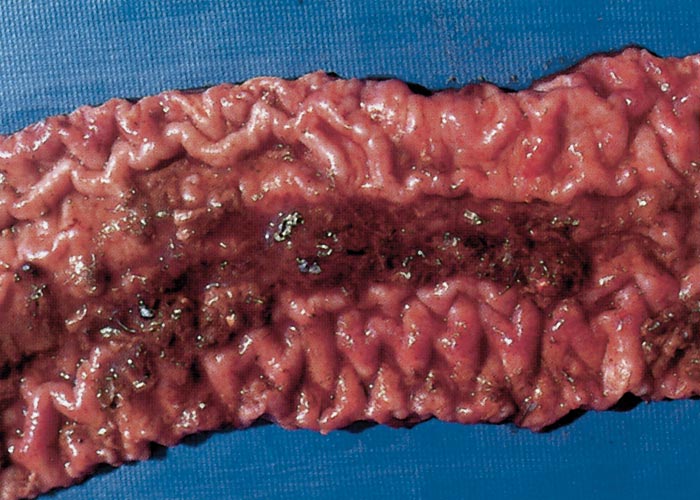
In pigs suffering from PHE that are killed when clinical signs first appear, there is marked oedema of the ileomesenteric attachment82, 113 and abundant clear ascitic fluid.82, 83 At this time the intestinal content is blood-tinged and the mucosa diffusely reddened (Figure 2). Twelve hours after the commencement of clinical signs, the mucosa is covered by a fibrinous pseudomembrane with discrete, fresh blood clots up to 10 mm in diameter trapped between the membrane and epithelium. Twenty-four hours after clinical signs have appeared, the lumen is filled with blood, and a cast consisting of clotted blood covers the mucosal surface.82, 83
The carcass shows generalized pallor in pigs that died of PHE.111-113, 120, 133 Specific lesions in most cases are confined to the ileum, but the caecum may also be affected in some.120 The serosa of the ileum has a characteristic reticulate appearance,82, 83, 120, 133 probably due to the subserosal oedema.133 The lumen of the ileum contains a large amount of fluid or clotted blood. The ileal mucosa, as in PIA, is thickened and ridged82, 83, 111-113, 120, 133 and pinkish-red to red. There may be mucosal petechiation.113, 133
Proliferation of immature mucosal epithelial cells, either as discrete islands within a normal mucosa or as diffuse involvement of the entire mucosa, is the underlying essential lesion common to PIA, NE, RI and PHE. This imparts the nodular or evenly thickened appearance to the intestine that can be observed macroscopically.21, 23, 71, 133 There is a sharp demarcation between normal and abnormal (adenomatous) mucosa.6, 21, 133 Villous atrophy causes a flattened mucosa,21, 132, 133 while the glands themselves are often highly distorted and branched.6, 23, 133
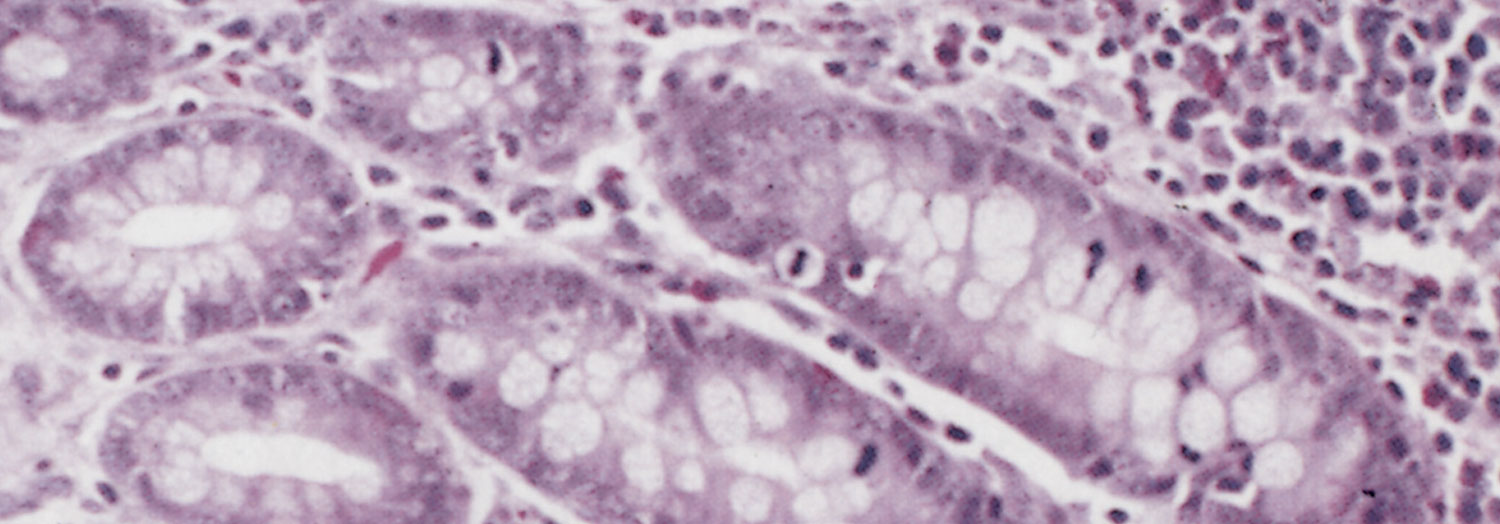
The glandular epithelial cells are immature, large, uniformly columnar, and often crowded onto the basal lamina to give an overlapping or pseudostratified appearance.6, 21, 23, 133 The cytoplasm is deeply eosinophilic to slightly basophilic.23 Most nuclei are either large and vesicular, containing one or two nucleoli, or elongated and densely stained.23, 133 Numerous cells undergoing mitosis6, 21, 23, 133 and an absence of goblet cells are evident in the adenomatous epithelium (compare Figure 3 and Figure 4).5 The marked immaturity of the proliferating epithelial cells is well demonstrated by positive pyroninophilia, poorly developed brush borders, and the slight cytoplasmic enzyme activity (alkaline phosphatase and adenosine triphosphatase). Taken as a whole, these staining reactions are typical of those of the undifferentiated epithelial cells at the base of mucosal glands.132, 133 Warthin-Starry silver impregnation techniques stain numerous, irregularly curved bacteria in the apical cytoplasm of the immature epithelial cells (Figure 5).71, 132 These intracellular bacteria can be identified as L. intracellularis with polyclonal,78 or more securely with monoclonal90 antibody in an indirect immunofluorescence test using formalin-fixed sections. As an alternative, the identity of the organism can be confirmed in frozen tissues by in situ hybridization using an L. intracellularis specific probe.32, 53
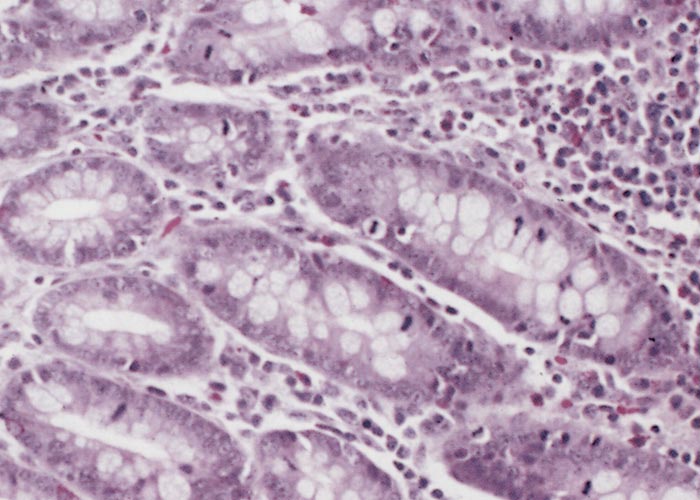
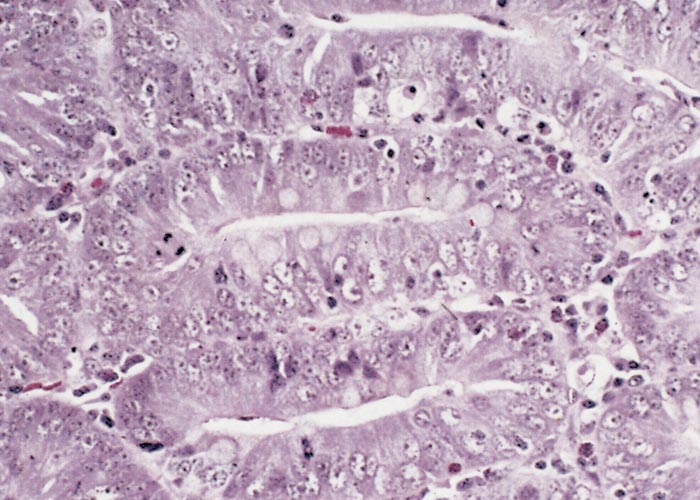
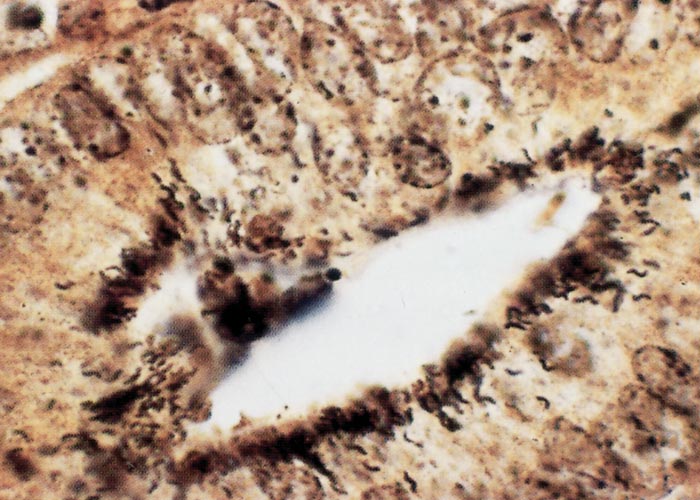
Changes in the intestinal wall (apart from those in the epithelium) are absent or mainly unremarkable in PIA. The exception is the occasional infiltration of diseased epithelium into the underlying tissue and spread of these cells to drainage lymph nodes.126 Increased numbers of T lymphocytes, plasma cells and a few eosinophils may be found in the lamina propria.21, 96, 133
In pigs killed immediately after the first appearance of clinical signs of PHE, histological evidence that may be related to an acute hypersensitivity reaction is found.82 This includes extensive accumulations of proteinaceous fluid in the interstitium and lacteals of the lamina propria, severe congestion of the mucosal blood vessels and marked infiltration of eosinophils into the mucosa. Mast cells and thrombosed vessels have been noted in the affected intestine.82
In pigs that have died of PHE, cell debris, either free in the lumen or attached to the mucosa, is invariably observed in addition to the adenomatous changes, blood and fibrin admixed with inflammatory cells.120, 133 Superficial epithelial desquamation111, 133 and necrosis of single or small groups133 of epithelial cells occur. Many mucosal glands are dilated and contain inflammatory cells, cell debris and desquamated epithelial cells.111, 120, 133 The lamina propria is usually moderately congested and oedematous.133 Haemorrhage is apparent in many areas in the lamina propria133 but this is not always seen.120 Histological evidence of blood vessel wall lesions is not observed.120, 133 Mild to moderate infiltration of inflammatory cells consisting of eosinophils, neutrophils, plasma cells and macrophages is usually present in the lamina propria.112, 120, 133 The submucosa is moderately to severely congested and oedematous,120 and may contain small clusters of neutrophils133 and a few eosinophils.133 Oedema of the intermuscular connective tissue in the tunica muscularis and of the subserosa has been described.133
Necrotic enteritis is characterized by patchy127, 129 to extensive6, 21 coagulative necrosis and inflammation of the mucosa. In some cases there is granulation tissue proliferation in the mucosa,129 while adenomatous epithelium may be found in non-necrotic areas of the mucosa.6, 127, 129
In RI the histological changes observed in the mucosa are variable. The mucosa may be adenomatous,23, 62, 119, 127, 129 chronically inflamed,23, 26, 119, 127, 129 or essentially normal.108 Typically, there is subacute inflammation of varying degrees with a mixed cell infiltrate in the lamina propria, submucosa and serosa.23, 62, 119, 129 This, together with marked hypertrophy of both layers of the tunica muscularis, is responsible for the thickening of the ileum observed macroscopically.23, 62, 108, 119, 129
Ultrastructurally, the epithelial cells in adenomatous areas of the mucosa resemble undifferentiated crypt cells.128 They are elongated, have a narrow apex,62, 128 and a basal nucleus62, 132 which is round, oval, or irregular and deeply indented.132 The cytoplasm contains numerous mitochondria of variable size,132 and clusters of free ribosomes. The few microvilli that are present are of irregular shape.62, 71, 128, 132 The lack of microvilli is more pronounced in cells that contain large numbers of intracytoplasmic bacteria.82
Intracellular bacteria are found in the immature cells in PIA,131 PHE,82 NE129 and RI.62 They are consistently found in the apical cytoplasm,62, 71, 128, 132 but in PHE bacteria also occur free, in macrophages in the lamina propria and submucosa, and within capillaries and lymphatics.82
Diagnosis
For a long time the diagnosis has been based on the demonstration of typical lesions at necropsy and by the use of staining techniques such as the Warthin-Starry impregnation of tissue sections of the lesions. However, these methods do not verify the identity of the microbe and small lesions may be overlooked. More recently, diagnostic methods such as PCR, immunohistochemistry and in situ hybridization have been developed.32, 53 Among these, identification of specific DNA by PCR and immunohistochemistry on tissue samples have been suggested as ‘gold standard’.63, 69
Confirmation of the diagnosis in clinically affected live animals can be obtained by the demonstration of L. intracellularis-specific DNA sequences by PCR in faecal samples. Initially, the diagnosis was based on a nested PCR.45, 61 This method has a good sensitivity and specificity but is time-consuming and thus expensive. More recently, the nested PCR has been replaced by fast, quantitative or semi-quantitative techniques based on real-time PCR, targeting the 16S rDNA or the aspA gene.81, 117 There has been a good correlation between the PCR results on faeces and the presence of histological PE lesions with intracellular bacteria.45, 53, 59 Clinical cases are likely to contain at least similar or greater numbers of L. intracellularis in their faeces.57
Lawsonia intracellularis can be detected in faeces using an indirect immunofluorescence test with either polyclonal or monoclonal antibody.139 However, the method has a rather poor sensitivity and requires the access to specific antibodies.
Several serological tests have been described for the detection of antibodies to L. intracellularis. These include a microscopic three-stage immunofluorescence test,76 and two tests using cultured bacteria: an immunofluorescence test67 and an immunoperoxidase monolayer assay. Several ELISAs have been described,41, 69, 152 one of which is commercially available.48
Using tissue-derived antigen, the presence of specific IgM antibodies correlates well with the presence of lesions but antibodies do not persist well, IgA and IgG titres being lower and sometimes absent in pigs that reveal lesions at necropsy.76 The use of cell-cultured antigen for detection of specific IgG appears more promising. In one investigation, most pigs exposed to infection appeared to seroconvert, but many of them did not appear to have developed lesions.67 Some studies indicated that circulating antibodies against L. intracellularis may be detected for five weeks only.38, 54 None of the tests can therefore be used to diagnose clinical cases with certainty and as with other tests they may only be available at specialist pig diagnostic laboratories.
Tissue culture has not been used successfully for the isolation of L. intracellularis from faeces.
A useful diagnostic aid for practitioners in the absence of laboratory help is the staining of mucosal smears from suspect lesions by the modified Ziehl-Neelsen technique.
Lawsonia intracellularis can be observed as clumps of red-stained, curved bacteria within blue epithelial cells.83 Although diagnosis in the individual animal may be important, the occurrence or absence of disease in a herd may be of more significance to the pig keeper. Examination of the intestines of abattoir pigs has been promoted as a means of detecting the disease,114 and this procedure may disclose disease previously undetected. Examination will often require to be supported by some of the abovementioned assays, as many thickened intestines are due to post-killing contraction.13, 40 The procedure may not reveal PE where the disease is mainly affecting post-weaned animals.40 There are, however, circumstances where an increase in cases in slaughter animals precedes the onset of clinical disease on the premises.114 Guidance for the selection and examination of intestines for PE has been published.115 However, this method is not used commonly these days.
Polymerase chain reaction examination of the faeces of post-weaned pigs will generally establish if the infection is present in the herd,12 and also may be used to establish the epidemiology of farm infection,46, 139 but may prove too expensive for this purpose. Although the PCR is likely to be reliably positive in clinical cases, some subclinical cases may be overlooked.139
Differential diagnosis
Clinically, PIA, NE and RI resemble a number of other enteric diseases of post-weaned pigs that manifest as diarrhoea and failure to make weight gains. There are also many specific and non-specific causes of poor weight gains in pigs. Most important of these are post-weaning multisystemic wasting syndrome (PMWS), chronic respiratory diseases, and spirochaetal or other forms of colitis. As many of the infections that can be confused with clinical and subclinical PE are also present in commercial pig herds, the relative importance of individual conditions may be difficult to assess.
The thickened ileal mucosal lesions of PIA are characteristic at necropsy. Post-weaning multisystemic wasting syndrome causes severe wasting in weaned pigs and in single cases, the disease may manifest as enteritis that macroscopically is indistinguishable from PE. However, the microscopic lesions differ and histiocytosis with the presence of PCV-2-positive cells in intestinal and lymphoid tissues distinguishes cases of PCV-2 enteritis.55 Swine dysentery and salmonellosis affect similar ages of pigs, but these conditions mainly affect the large bowel and the necrotic lesions are more superficial. Salmonellosis is generally a febrile disease. Clostridium perfringens Type C infection and coccidiosis can cause necrotic enteritis, but usually affect pigs outside the post-weaned age group.
The characteristic melaena of clinical PHE may be confused with other diseases that manifest the same clinical signs, namely gastric ulcer, intestinal volvulus, warfarin toxicosis, swine dysentery, sporadic cases of Escherichia coli serotype O149K88 (F4), Salmonella cholerasuis infection, and intestinal anthrax. At necropsy, PHE cases must be distinguished from a sporadic, but common, intestinal catastrophe described as colonic bloat, intestinal torsion, intestinal haemorrhage syndrome or haemorrhagic bowel syndrome.142
Control
The need to control PHE is obvious as the disease in a susceptible herd leads to significant economic losses, especially where the animals are derived from high health status stock. The losses due to PIA, however, may be of greater consequence but are often unrealized. The loss caused by infection has been assessed at between UK £2 and £7 per affected pig.102 The widespread distribution of the disease should encourage producers to consider the potential impact of infection.
One of the most disturbing aspects of the infection is the appearance of disease in many recently established high health status herds, either derived from medicated early-weaned or hysterectomy-derived progeny, or their descendants.140 Unfortunately, no specific recommendations can be provided that will absolutely minimize this risk, as the source of infection often remains unclear.
Proliferative enteropathy can be treated with a variety of antibacterials of which the most often used are chlortetracycline, doxycycline, tiamulin or tylosin.70, 99, 101, 103 It should be noted that L. intracellularis has recently been shown to carry a gene that might confer resistance to tetracycline.147 Results of treatment of clinical PHE are poor irrespective of the antibiotic used.83, 120, 133, 134 To comply with a prudent use of antibiotics, the best strategy for treatment should be carefully considered. In cases of single ill pigs in a group of otherwise healthy animals, individual treatments should be preferred. To minimize the antibiotic usage, diseased pigs could preferably be gathered, isolated and treated in a separate pen. If extensive problems occur, batch-wise medication could be considered but should always be combined with other measures. The use of in-feed or water medication should be limited to a certain time-period or a certain batch of pigs, and should be accompanied by the implementation of all-in all-out and batch-wise production systems including thorough cleaning and disinfection between each batch, to minimize the impact of residual bacteria.15 The bacteria may form a biofilm147 but are sensitive to several disinfectants, are completely inactivated by quaternary ammonium compounds, and will be inactivated to ≥99 per cent by oxidising agents.153
At present, one vaccine is available on the market.110 It is a live attenuated vaccine that can be administered by oral drench to the piglets or in water to, for example, finisher pigs. The vaccine cannot be given concurrently with the use of antibiotics and a minimum of one week medication-free “window” is required, with vaccination taking place on the fourth day. New vaccines are currently under development. At farm level, it is advisable to establish the time-point for the onset of the infection and administer the vaccine at least three weeks ahead, or before the pigs are being moved to the actual facilities.150 In the recruitment of pigs from high-health herds that will enter a herd known to be infected, it is advisable to vaccinate the recruitment animals in quarantine, three weeks before they are allowed to enter the farm.
References
- ALBERDI, M. P., WATSON, E., MCALLISTER, G. E. M., HARRIS, J. D., PAXTON, E. A., THOMSON, J. R. & SMITH, D. G. E., 2009. Expression by Lawsonia intracellularis of type III secretion system components during infection. Veterinary Microbiology, 139, 298-303.
- ALDERTON, M. R., BORLAND, R. & COLOE, P. J., 1992. Experimental reproduction of porcine proliferative enteritis. Journal of Comparative Pathology, 106, 159–167.
- BACKHANS, A., JACOBSON, M., HANSSON, I., LEBBAD, M., THISTED LAMBERTZ, S., GAMMELGARD, E., SAAGER, M., AKANDE, O. & FELLSTRÖM, C., 2013. Occurrence of pathogens in wild rodents caught on Swedish pig and chicken farms. Epidemiology and Infection, 141, 1885-1891.
- BEERS, P. T., 1984. Studies on porcine adenomatosis with particular reference to haemorrhagic enteropathy. PhD thesis, Sydney University, Australia.
- BENGTSSON, R. J., MACINTYRE, N., GUTHRIE, J., WILSON, A. D., FINLAYSON, H., MATIKA, O., PONG-WON, R., SMITH, S. H., ARCHIBALD, A. L. & AIT-ALI, T., 2015. Lawsonia intracellularis infection of intestinal crypt cells is associated with specific depletion of secreted MUC2 in goblet cells. Veterinary Immunology and Immunopathology, 168, 61-67.
- BIESTER, H. E. & SCHWARTE, L. H., 1931. Intestinal adenoma in swine. American Journal of Pathology, 7, 175-185.
- BIESTER, H. E., SCHWARTE, L. H. & EVELETH, D. F., 1939. Studies on a rapidly developing intestinal adenoma in a pig. American Journal of Pathology, 15, 385–389.
- BOESEN, H. T., JENSEN, T. K., SCHMIDT, A. S., JENSEN, B. B., JENSEN, S. M. & MÖLLER, K., 2004. The influence of diet on Lawsonia intracellularis colonization in pigs upon experimental challenge. Veterinary Microbiology, 103, 35-45.
- BOOSINGER, T. R., THACKER, H. L. & ARMSTRONG, C. H., 1985. Campylobacter sputorum subspecies mucosalis and Campylobacter hyointestinalis infections in the intestine of gnotobiotic pigs. American Journal of Veterinary Research, 46, 2152–2156.
- BOUTRUP, T. S., BOESEN, H. T., BOYE, M., AGERHOLM, J. S. & JENSEN, T. K., 2010. Early pathogenesis in porcine proliferative enteropathy caused by Lawsonia intracellularis. Journal of Comparative Pathology, 143, 101-109.
- BOUTRUP, T. S., SCHAUSER, K., AGERHOLM, J. S. & JENSEN, T. K., 2010. Application of a pig ligated intestinal loop model for early Lawsonia intracellularis infection. Acta Veterinaria Scandinavica, 52, 1-7. http://www.actavetscand.com/content/52/1/17.
- CHANG, W. L., WU, C. F., WU, Y., KAO, Y. M. & PAN, M. J., 1997. Prevalence of Lawsonia intracellularis in swine herds in Taiwan. The Veterinary Record, 141, 103-104.
- CHRISTENSEN, N. H. & CULLINANE, L. C., 1990. Monitoring the health of pigs in New Zealand abattoirs. New Zealand Veterinary Journal, 38, 136–141.
- COLLINS, A. M. & BARCHIA, I. M., 2014. The critical threshold of Lawsonia intracellularis in pig faeces that causes reduced average daily weight gains in experimentally challenged pigs. Veterinary Microbiology, 168, 455-458.
- COLLINS, A. M., FELL, S. A. & BARCHIA, I. M., 2013. Cleaning and disinfection with Virkon S significantly reduces Lawsonia intracellularis survival and transmission to naive pigs. Journal of Swine Health and Production, 21, 144-147.
- COLLINS, A. M., SWIFT, I. & MONKTON, R. P., 1996. Replication of Australian porcine isolates of ileal symbiont intracellularis in tissue culture. Veterinary Microbiology, 49, 249–255.
- COOPER, D. M., SWANSON, D. L., BARNS, S. M. & GEBHART, C. J., 1997. Comparison of the 16S ribosomal DNA sequence from the intracellular agent of proliferative enteritis in a hamster, deer, and ostrich with the sequence of a porcine isolate of Lawsonia intracellularis. International Journal of Systematic Bacteriology, 47, 635–639.
- CORDES, H., RIBER, U., JENSEN, T. K. & JUNGERSEN, G., 2012. Cell-mediated and humoral immune responses in pigs following primary and challenge-exposure to Lawsonia intracellularis. Veterinary Research, 43, 1-11. http://www.veterinaryresearch.org/content/43/1/9.
- CROSS, R. F., SMITH, C. K. & PARKER, C. F., 1973. Terminal ileitis in lambs. Journal of the American Veterinary Medical Association, 162, 564–566.
- DALE, C. J. H., MOSES, E. K., ONG, C. C., MORROW, C. J., REED, M. B., HASSE, D. & STRUGNELL, R. A., 1998. Identification and sequencing of the groE operon and flanking genes of Lawsonia intracellularis: use in phylogeny. Microbiology, 144, 2073–2084.
- DODD, D. C., 1968. Adenomatous intestinal hyperplasia (proliferative enteritis) of swine. Pathologia Veterinaria, 5, 333–342.
- DUHAMEL, G. E. & WHEELDON, E. B., 1982. Intestinal adenomatosis in a foal. Veterinary Pathology, 19, 447–450.
- EMSBO, P., 1951. Terminal or regional ileitis in swine. Nordisk Veterinaer Medicin, 3, 1-28.
- ERIKSEN, K., LANDSVERK, T. & BRATBERG, B., 1990. Morphology and immunoperoxidase studies of intestinal adenomatosis in the blue fox, Alopex lagopus. Journal of Comparative Pathology, 102, 265–278.
- ERIKSEN, K., LANDSVERK, T., GONDROSEN, B. & VORMELAND, J., 1990. Immunohistochemical and cytochemical evidence suggesting the presence of Campylobacter jejuni and Campylobacter coli in cases of porcine intestinal adenomatosis. Acta Veterinaria Scandinavica, 31, 445–451.
- FIELD, H. L., BUNTAIN, D. & JENNINGS, A. R., 1953. Terminal or regional ileitis in pigs. Journal of Comparative Pathology, 63, 153–159.
- FOX, J. G., MURPHY, J. C., ACKERMAN, J. I., PROSTAK, K. S., GALLAGHER, C. A. & RAMBOW, V. J., 1982. Proliferative colitis in ferrets. American Journal of Veterinary Research, 43, 858–864.
- FRIEDMAN, M., BEDNÁL, V., KLIMEŠ, J., SMOLA, J., MRLIK, V. & LITERÁK, I., 2008. Lawsonia intracellularis in rodents from pig farms with the occurrence of porcine proliferative enteropathy. Letters in Applied Microbiology, 47, 117-121.
- FRIENDSHIP, C. R. & BILKEI, G., 2007. Concurrent swine erysipelas and Clostridium novyi infections associated with sow mortality in outdoor sows in Kenya. Veterinary Journal, 173, 694-696.
- GEBHART, C. J., BARNS, S. M., MCORIST, S., LIN, G. F. & LAWSON, G. H. K., 1993. Ileal symbiont intracellularis, an obligate bacterium of porcine intestine showing a relationship to Desulfovibrio species. International Journal of Systematic Bacteriology, 43, 533–538.
- GEBHART, C. J., KURTZ, H. J., WARD, G. E. & CHANG, K., 1985. The hamster as a model for porcine proliferative enteritis. In: PEARSON, A. D., SKIRROW, M. B., LIOR, H. & ROWE, B., (eds.). Campylobacter III. Proceedings of the Third International Workshop on Campylobacter Infections. Public Health Laboratory Service, London, U.K. 99.
- GEBHART, C. J., MCORIST, S., LAWSON, G. H. K., COLLINS, J. E. & WARD, G. E., 1994. Specific in situ hybridisation of the intracellular organism of the porcine proliferative enteropathy. Veterinary Pathology, 31, 462–467.
- GEBHART, C. J., WARD, G. E., CHANG, K. & KURTZ, H. J., 1983. Campylobacter hyointestinalis (new species) isolated from swine with lesions of proliferative enteritis. American Journal of Veterinary Research, 44, 361–367.
- GOGOLEWSKI, R. P., COOK, R. W. & BATTERHAM, E. S., 1991. Suboptimal growth associated with porcine intestinal adenomatosis in pigs in nutritional studies. Australian Veterinary Journal, 68, 406-408.
- GUEDES, R. M. C. & GEBHART, C. J., 2003a. Comparison of intestinal mucosa homogenate and pure culture of the homologous Lawsonia intracellularis isolate in producing proliferative enteropathy in swine. Veterinary Microbiology, 93, 159-166.
- GUEDES, R. M. C. & GEBHART, C. J., 2003b. Onset and duration of fecal shedding, cell-mediated and humoral immune responses in pigs after challenge with a pathogenic isolate or attenuated vaccine strain of Lawsonia intracellularis. Veterinary Microbiology, 91, 135-145.
- GUEDES, R. M. C. & GEBHART, C. J., 2010. Evidence of cell-mediated immune response and specific local mucosal immunoglobulin (Ig) QA production against Lawsonia intracellularis in experimentally infected swine. Canadian Journal of Veterinary Research, 74, 97-101.
- GUEDES, R. M. C., GEBHART, C. J., ARMBRUSTER, G. A. & ROGGOW, B. D., 2002. Serologic follow-up of a repopulated swine herd after an outbreak of proliferative hemorrhagic enteropathy. Canadian Journal of Veterinary Research, 66, 258-263.
- GUEDES, R. M. C., WINKELMAN, N. L. & GEBHART, C. J., 2003. Relationship between the severity of porcine proliferative enteropathy and the infectious dose of Lawsonia intracellularis. Veterinary Record, 153, 432-433.
- HOLYOAKE, P. K., CUTLER, R. S. & CAPLE, I. W., 1994. A diagnostic dilemma: detecting proliferative enteropathy in pigs at slaughter. Australian Veterinary Journal, 71, 308–309.
- HOLYOAKE, P. K., CUTLER, R. S., CAPLE, I. W. & MONKTON, R. P., 1994. Enzyme-linked immunosorbent assay for measuring Ileal symbiont intracellularis specific immunoglobulin G response in the sera of pigs. Journal of Clinical Microbiology, 32(8), 1980–1985.
- HOLYOAKE, P. K., EMERY, D., GONSALVES, J., DONAHOO, M. & COLLINS, A. M., 2010. Prevalence of antibodies to Lawsonia intracellularis in pig herds in Australia. Australian veterinary journal, 88, 186-188.
- JACKSON, G. H. & BAKER, J. R., 1980. Occurrence of unthriftiness in piglets post-weaning. Pig Veterinary Society Proceedings, 7, 63–67.
- JACOBSON, M., ANDERSSON, M., LINDBERG, R., JENSEN-WAERN, M. & FOSSUM, C., 2011. Microarray and cytokine analyses of field cases of pigs with diarrhoea. Veterinary Microbiology, 153, 307-314.
- JACOBSON, M., ASPAN, A., HELDTANDER KÖNIGSSON, M., HARD, A. F., SEGERSTAD, C., WALLGREN, P., FELLSTRÖM, C., JENSEN-WAERN, M. & GUNNARSSON, A., 2004. Routine diagnostics of Lawsonia intracellularis performed by PCR, serological and post mortem examination, with special emphasis on sample preparation methods for PCR. Veterinary Microbiology, 102, 189–201.
- JACOBSON, M., ASPAN, A., NORDENGRAHN, A., LINDBERG, M. & WALLGREN, P., 2010. Monitoring of Lawsonia intracellularis in breeding herd gilts. Veterinary Microbiology, 142, 317-322.
- JACOBSON, M., GERTH LÖFSTEDT, M., HOLMGREN, N., LUNDEHEIM, N. & FELLSTRÖM, C., 2005. The prevalence of Brachyspira spp.and Lawsonia intracellularis in Swedish swine herds and in the wild boar population. Journal of Veterinary Medicine series B, 52, 386-391.
- JACOBSON, M., WALLGREN, P., NORDENGRAHN, A., MERZA, M. & EMANUELSON, U., 2011. Evaluation of a blocking ELISA for the detection of antibodies to Lawsonia intracellularis in pig sera. Acta Veterinaria Scandinavica 53, 1-6. http://www.actavetscand.com/content/53/1/23.
- JACOBY, R. O., 1978. Transmissible ileal hyperplasia of hamsters. American Journal of Pathology, 91, 433–444.
- JACOBY, R. O., OSBALDISTON, G. W. & JONAS, A. W., 1975. Experimental transmission of atypical ileal hyperplasia of hamsters. Laboratory Animal Science, 25, 465–473.
- JASNI, S., MCORIST, S. & LAWSON, G. H. K., 1994. Reproduction of proliferative enteritis in hamsters with a pure culture of porcine Ileal symbiont intracellularis. Veterinary Microbiology, 41, 1-9.
- JENSEN, T. K., CHRISTENSEN, B. B. & BOYE, M., 2006. Lawsonia intracellularis infection in the large intestines of pigs. Acta Pathologica, Microbiologica et Immunologica Scandinavica, 114, 255–264.
- JENSEN, T. K., MOLLER, K., LESER, T. D. & JORSAL, S. E., 1997. Comparison of histology, immunohistochemistry and polymerase chain reaction for detection of Lawsonia intracellularis in natural porcine proliferative enteropathy. European Journal of Veterinary Pathology, 3, 115-123.
- JENSEN, T. K., VIGRE, H., SÖRENSEN, V. & MÖLLER, K., 2005. Naturally acquired Lawsonia intracellularis infection in pigs studied from weaning to slaughter by indirect immunofluorescence antibody test and polymerase chain reaction on faeces. Research in Veterinary Science, 79, 93–98.
- JENSEN, T. K., VIGRE, H., SVENSMARK, B. & BILLE-HANSEN, V., 2006. Distinction between porcine circovirus type 2 enteritis and porcine proliferative enteropathy caused by Lawsonia intracellularis. Journal of Comparative Pathology, 135, 176-182.
- JOENS, L. A., NIBBELINK, S. & GLOCK, R. D., 1997. Induction of gross and microscopic lesions of porcine proliferative enteritis by Lawsonia intracellularis. American Journal of Veterinary Research, 58, 1125–1131.
- JOHANSEN, M., NIELSEN, M., DAHL, J., SVENSMARK, B., BAEKBO, P., KRISTENSEN, C. S., HJULSAGER, C. K., JENSEN, T. K., STAHL, M., LARSEN, L. E. & ANGEN, O., 2013. Investigation of the association of growth rate in grower-finisher pigs with the quantification of Lawsonia intracellularis and porcine circovirus type 2. Preventive Veterinary Medicine, 63-72.
- JOHNSON, E. A. & JACOBY, R. O., 1978. Transmissible ileal hyperplasia of hamsters. American Journal of Pathology, 91, 451–459.
- JONES, G. F., DAVIES, P. R., ROSE, R., WARD, G. E. & MURTAUGH, M. P., 1993. Comparison of techniques for diagnosis of proliferative enteritis of swine. American Journal of Veterinary Research, 54, 1980–1985.
- JONES, G. F., WARD, G. E., COLLINS, J. E. & GEBHART, C. J., 1993. Transmission of proliferative enteritis to swine by use of embryonated eggs. American Journal of Veterinary Research, 54, 1256–1261.
- JONES, G. F., WARD, G. E., MURTAUGH, M. P., LIN, G. & GEBHART, C. J., 1993. Enhanced detection of intracellular organism of swine proliferative enteritis, Ileal symbiont intracellularis, in feces by polymerase chain reaction. Journal of Clinical Microbiology, 31, 2611–2615.
- JONSSON, L. & MARTINSSON, K., 1976. Regional ileitis in pigs. Morphological and pathogenetical aspects. Acta Veterinaria Scandinavica, 17, 223–232.
- JORDAN, D. M., KNITTEL, J. P., ROOF, M. B., SCHWARTZ, K., LARSON, D. & HOFFMAN, L. J., 1999. Detection of Lawsonia intracellularis in swine using polymerase chain reaction methodology. Journal of Veterinary Diagnostic Investigation, 11, 45-49.
- JUST, S. D., THOEN, C. O., THACKER, B. J. & THOMPSON, J. U., 2000. Monitoring of Lawsonia intracellularis by indirect serum immunofluorescence assay in a commercial swine production system. Journal of Swine Health and Production, 9, 57-61.
- KIM, J., WON, G., PARK, S. & LEE, J. H., 2017. Identification of Lawsonia intracellularis putative hemolysin protein A and characterization of its immunoreactivity. Veterinary Microbiology, 205, 57-61.
- KIM, O., KIM, B. & CHAE, C., 1998. Prevalence of Lawsonia intracellularis in selected pig herds in Korea as determined by PCR. The Veterinary Record, 587-589.
- KNITTEL, J. P., JORDAN, D. M., SCHWARTZ, K. J., JANKE, B. H., ROOF, M. B., MCORIST, S. & HARRIS, D. L., 1998. Evaluation of antemortem polymerase chain reaction and serologic methods for detection of Lawsonia intracellularis-exposed pigs. American Journal of Veterinary Research, 59, 722–726.
- KNITTEL, J. P., LARSON, D. I., HARRIS, D. L., ROOF, M. B. & MCORIST, S., 1996. United States isolates of Lawsonia intracellularis from porcine proliferative enteropathy resemble European isolates. Swine Health and Production, 4, 119–122.
- KROLL, J. J., EICHMEYER, M. A., SCHAEFFER, M. L., MCORIST, S., HARRIS, D. L. & ROOF, M. B., 2005. Lipopolysaccharide-based enzyme-linked immunosorbent assay for experimental use in detection of antibodies to Lawsonia intracellularis in pigs. Clinical and Diagnostic Laboratory Immunology, 12, 693–699.
- KYRIAKIS, S. C., BOURTZI-HATZOPOULOU, E., ALEXOPOULOS, C., KRITAS, K., POLYZOPOULOU, Z., LEKKAS, S. & GARDEY, L., 2002. Field evaluation of the effect of in-feed doxycycline for the control of ileitis in weaned piglets. Journal of Veterinary Medicine series B, 49, 317-321.
- LANDSVERK, T. & NORDSTOGA, K., 1981. Intestinal adenomatosis in pigs. A patho-morphological investigation. Nordisk Veterinaer Medicin, 36, 77-80.
- LAWSON, G. H. K. & GEBHART, C. J., 2000. Proliferative enteropathy. Journal of Comparative Pathology, 122, 77-100.
- LAWSON, G. H. K., MACKIE, R. A. M., SMITH, D. G. E. & MCORIST, S., 1995. Infection of cultured rat enterocytes by Ileal symbiont intracellularis depends on host cell function and actin polymerisation. Veterinary Microbiology, 45, 339–350.
- LAWSON, G. H. K. & MCORIST, S., 1993. The enigma of the proliferative enteropathies: A review. Journal of Comparative Pathology, 108, 41-46.
- LAWSON, G. H. K., MCORIST, S., JASNI, S. & MACKIE, R. A., 1993. Intracellular bacteria of porcine proliferative enteropathy: Cultivation and maintenance in vitro. Journal of Clinical Microbiology, 31, 1136–1142.
- LAWSON, G. H. K., MCORIST, S., ROWLAND, A. C., MCCARTNEY, E. & ROBERTS, L., 1988. Serological diagnosis of the porcine proliferative enteropathies: Implications for aetiology and epidemiology. The Veterinary Record, 122, 554–557.
- LAWSON, G. H. K. & ROWLAND, A. C., 1994. Intestinal adenomatosis in the pig: A bacteriological study. Research in Veterinary Science, 17, 331-336.
- LAWSON, G. H. K., ROWLAND, A. C. & MACINTYRE, N., 1985. Demonstration of a new intracellular antigen in porcine intestinal adenomatosis and hamster proliferative ileitis. Veterinary Microbiology, 10, 303–313.
- LAWSON, G. H. K., ROWLAND, A. C., ROBERTS, L., FRASER, G. & MCCARTNEY, E., 1979. Proliferative haemorrhagic enteropathy. Research in Veterinary Science, 27, 46-51.
- LAWSON, G. H. K., ROWLAND, A. C., SMITH, W. J., ROBERTS, L. & LUNNY, D., 1980. Immunisation of pigs with Campylobacter sputorum subspecies mucosalis vaccine. The Veterinary Record, 107, 424–425.
- LINDECRONA, R. H., JENSEN, T. K., ANDERSEN, P. H. & MOLLER, K., 2002. Application of a 5’ nuclease assay for detection of Lawsonia intracellularis in fecal samples from pigs. Journal of Clinical Microbiology, 40, 984-987.
- LOVE, D. N. & LOVE, R. J., 1979. Pathology of proliferative haemorrhagic enteropathy. Veterinary Pathology, 16, 41–48.
- LOVE, D. N., LOVE, R. J. & EDWARDS, M. J., 1977. Proliferative haemorrhagic enteropathy in pigs. The Veterinary Record, 100, 65–68.
- MAPOTHER, M. E., JOENS, L. A. & GLOCK, R. D., 1987. Experimental reproduction of porcine proliferative enteritis. The Veterinary Record, 121, 533-536.
- MAPOTHER, M. E., JOENS, L. A. & GLOCK, R. D., 1987. Investigations into the aetiology of porcine proliferative enteritis. The Veterinary Record, 121, 86.
- MARTINSSON, K., EKMAN, L. & JONSSON, L., 1976. Haematological and biochemical analysis of blood and serum in pigs with regional ileitis with special reference to the pathogenesis.,em>. Acta Veterinaria Scandinavica, 17, 233–243.
- MCCARTNEY, E., LAWSON, G. H. K. & ROWLAND, A. C., 1984. Behaviour of Campylobacter sputorum subspecies mucosalis in gnotobiotic pigs. Research in Veterinary Science, 36, 290-297.
- MCCLUSKEY, J., HANNIGAN, J., HARRIS, J. D., WREN, B. & SMITH, D. G. E., 2002. LsaA, an antigen involved in cell attachment and invasion, is expressed by Lawsonia intracellularis during infection in vitro and in vivo. Infection and Immunity, 70, 2899-2907.
- MCORIST, S., BOID, R. & LAWSON, G. H. K., 1989. Antigenic analysis of Campylobacter species and an intracellular Campylobacter-like organism associated with porcine proliferative enteropathies. Infection and Immunity, 57, 957–962.
- MCORIST, S., BOID, R., LAWSON, G. H. K. & MCCONNELL, I., 1987. Monoclonal antibodies to intracellular Campylobacter-like organisms of the porcine proliferative enteropathies. The Veterinary Record, 121, 421–422.
- MCORIST, S., GEBHART, C. J., BOID, R. & BARNS, S. M., 1995. Characterization of Lawsonia intracellularis gen. nov., sp. nov., the obligate intracellular bacterium of porcine proliferative enteropathy. International Journal of Systematic Bacteriology, 45, 820–825.
- MCORIST, S., JASNI, S., MACKIE, R. A., MACINTYRE, N., NEEF, N. & LAWSON, G. H. K., 1993. Reproduction of porcine proliferative enteropathy with pure cultures of Ileal symbiont intracellularis. Infection and Immunity, 61, 4286–4292.
- MCORIST, S., JASNI, S., MACKIE, R. A. M., BERSCHNEIDER, H. M., ROWLAND, A. C. & LAWSON, G. H. K., 1995. Entry of the bacterium Ileal symbiont intracellularis into cultured enterocytes and its subsequent release. 59, Research in Veterinary Science, 255–260.
- MCORIST, S. & LAWSON, G. H. K., 1989. Reproduction of proliferative enteritis in gnotobiotic pigs. Research in Veterinary Science, 46, 27-33.
- MCORIST, S., LAWSON, G. H. K., ROWLAND, A. C. & MACINTYRE, N., 1989. Early lesions of proliferative enteritis in pigs and hamsters. Veterinary Pathology, 26, 260-264.
- MCORIST, S., MACINTYRE, N., STOKES, C. R. & LAWSON, G. H. K., 1992. Immunocytological responses in porcine proliferative enteropathies. Infection and Immunity, 60, 4184–4191.
- MCORIST, S., MACKIE, R. A. M., LAWSON, G. H. K. & SMITH, D. G. E., 1997. In-vitro interactions of Lawsonia intracellularis with cultured enterocytes. Veterinary Microbiology, 54, 385–392.
- MCORIST, S., MACKIE, R. A. M., NEEF, N., AITKEN, I. & LAWSON, G. H. K., 1994. Synergism of ileal symbiont intracellularis and gut bacteria in the reproduction of porcine proliferative enteropathy. The Veterinary Record, 134, 331–332.
- MCORIST, S., MORGAN, J. H., VEENHUIZEN, M., LAWRENCE, K. & KROGER, H. W., 1997. Oral administration of tylosin phosphate and prevention of proliferative enteropathy in pigs. American Journal of Veterinary Research, 58, 136-139.
- MCORIST, S., ROBERTS, L., JASNI, S., ROWLAND, A. C., LAWSON, G. H. K., GEBHART, C. J. & BOSWORTH, B., 1996. Developed and resolving lesions in porcine proliferative enteropathy: Possible pathogenic mechanisms. (1996). Journal of Comparative Pathology, 115, 35-45.
- MCORIST, S., SHEARN, M. F. H. & MORGAN, J. D., 1999. Control of porcine proliferative enteropathy by oral administration of chlortetracycline. The Veterinary Record, 144, 48-49.
- MCORIST, S., SMITH, S. H. & GREEN, L. E., 1997. Estimate of direct financial losses due to porcine proliferative enteropathy. The Veterinary Record, 140, 579–581.
- MCORIST, S., SMITH, S. H., SHEARN, M. F. H., CARR, M. M. & MILLER, D. J. S., 1996. Treatment and prevention of proliferative enteropathy with oral tiamulin. The Veterinary Record, 139, 615–618.
- MIRAJKAR, N. S., KELLEY, M. R. & GEBHART, C. J., 2017. Draft genome sequence of Lawsonia intracellularis strain E40504, isolated from a horse diagnosed with equine proliferative enteropathy. Genome Announcements, 5, e00330-17. https://doi.org/10.1128/genomeA00330-17.
- MOLLER, K., JENSEN, T. K. & JORSAL, S. E., 1998. Detection of Lawsonia intracellularis in endemically infected pig herds. Proceedings of the International Pig Veterinary Society, 15th Congress, 2, 63.
- MOLLER, K., JENSEN, T. K., JORSAL, S. E., LESER, T. D. & CARSTENSEN, B., 1998. Detection of Lawsonia intracellularis, weakly beta-haemolytic intestinal spirochaetes, Salmonella enterica, and haemolytic Escherichia coli from swine herds with and without diarrhoea among growing pigs. Veterinary Microbiology, 62, 59-72.
- MOON, H. W., CUTLIP, R. C., AMTOWER, W. C. & MATTHEWS, P. J., 1974. Intraepithelial vibrio associated with acute typhilitis of young rabbits. Veterinary Pathology, 11, 313–326.
- NIELSEN, S. W., 1955. Muscular hypertrophy of the ileum in relation to terminal ileitis in pigs—a preliminary report. Journal of the American Veterinary Medical Association, 36, 290–297.
- NISHIKAWA, S., OGAWA, Y., EGUCHI, M., RAMBUKKANA, A. & SHIMOJI, Y., 2018. Draft genome sequences of Lawsonia intracellularis swine strains causing proliferative enteropathy in Japan. Microbiology Resource Announcements, 7, e01021-18. https://doi.org/10.1128/MRA.01021-18.
- NOGUEIRA, M. G., COLLINS, A. M., DONAHOO, M. & EMERY, D., 2013. Immunological responses to vaccination following experimental Lawsonia intracellularis virulent challenge in pigs. Veterinary Microbiology, 164, 131-138.
- O’HARA, P. J., 1972. Intestinal haemorrhage syndrome in the pig. Veterinary Record, 91, 517–518.
- O’NEILL, P. A., 1970. Observations on a haemorrhagic bowel syndrome involving pigs on three associated premises. Veterinary Record, 87, 742–747.
- PILL, A. H., 1971. A haemorrhagic bowel syndrome in pigs. Veterinary Record, 87, 27–28.
- POINTIN, A. M., 1989. Campylobacter associated intestinal pathology in pigs. Australian veterinary journal, 66, 90-91.
- POINTIN, A. M., MERCY, A. R., BACKSTROM, L. & DIAL, G. D., 1992. Disease surveillance at slaughter. In: LEMAN, A. D., STRAW, B. E., MENGELING, W. L., D’ALLAIRE, S. & TAYLOR, D. J.,(eds.). Diseases of Swine. 7th edn. Ames, Iowa: Iowa State University Press.
- POZO, J., CARVAJAL, A., DE ARRIBA, M. L., LANZA, I. & RUBIO, P., 1998. Presence of Lawsonia intracellularis as an aetiological agent in enteric disorders in Spanish swine herds. Proceedings of the International Pig Veterinary Society, 15th Congress, 3, 416.
- PUSTERLA, N., MAPES, S., REJMANEK, D. & GEBHART, C. J., 2008. Detection of Lawsonia intracellularis by real-time PCR in the feces of free-living animals from equine farms with documented occurrence of equine proliferative enteropathy. Journal of Wildlife Diseases, 44, 992-998.
- PUSTERLA, N., SANCHEZ-MIGALLON GUZMAN, D., VANNUCCI, F. A., MAPES, S., WHITE, A., DIFRANCESCO, M. & GEBHART, C. J., 2013. Transmission of Lawsonia intracellularis to weanling foals using feces from experimentally infected rabbits. Veterinary Journal, 195, 241-243.
- RAKHO, T. & SALONIEMI, H., 1972. On the pathology of regional ileitis in the pig. Nordisk Veterinaer Medicin, 24, 132-138.
- REDMAN CHU, R. M. & HONG, C. B., 1973. Haemorrhagic bowel syndrome in pigs in Taiwan. The Veterinary Record, 93, 562.
- RESENDE, T. P., REAL PEREIRA, C. E., DE SOUZA DANIEL, A. G., VASQUEZ, E., SAQUI-SALCES, M., VANUCCI, F. A. & GEBHART, C. J., 2018. Effects of Lawsonia intracellularis infection in the proliferation of different mammalian cell lines. Veterinary Microbiology, 228, 157-164.
- ROBERTS, L., LAWSON, G. H. K. & ROWLAND, A. C., 1980. The experimental infection of neonatal pigs with Campylobacter sputorum subsp. mucosalis. Research in Veterinary Science, 23, 257–258.
- ROBERTS, L., LAWSON, G. H. K. & ROWLAND, A. C., 1980. The experimental infection of pigs with Campylobacter sputorum subsp. mucosalis. Weaned pigs with special reference to pharmacologically mediated hypomotility. Research in Veterinary Science, 28, 148-150.
- ROBERTS, L., LAWSON, G. H. K., ROWLAND, A. C. & LAING, A. H., 1979. Porcine intestinal adenomatosis and its detection in a closed pig herd. The Veterinary Record, 104, 366–368.
- ROBERTS, L., ROWLAND, A. C. & LAWSON, G. H. K., 1977. Experimental reproduction of porcine intestinal adenomatosis and necrotic enteritis. The Veterinary Record, 100, 12-13.
- ROBERTS, L., ROWLAND, A. C. & LAWSON, G. H. K., 1980. Porcine intestinal adenomatosis: epithelial dysplasia and infiltration. Gut, 21, 1035–1040.
- ROWLAND, A. C. & HUTHCHINGS, D. A., 1978. Necrotic enteritis and regional ileitis in pigs at slaughter. The Veterinary Record, 103, 338–339.
- ROWLAND, A. C. & LAWSON, G. H. K., 1974. Intestinal adenomatosis in the pig: immunofluorescent and electron microscopic studies. Research in Veterinary Science, 17, 323–330.
- ROWLAND, A. C. & LAWSON, G. H. K., 1975. Porcine intestinal adenomatosis: A possible relationship with necrotic enteritis, regional ileitis and proliferative haemorrhagic enteropathy. The Veterinary Record, 97, 178–180.
- ROWLAND, A. C. & LAWSON, G. H. K., 1975b. Intestinal adenomatosis in the pig: a possible relationship with a haemorrhagic enteropathy. Research in Veterinary Science, 18, 263-268.
- ROWLAND, A. C., LAWSON, G. H. K. & MAXWELL, A., 1973. Intestinal adenomatosis in the pig: Occurrence of a bacterium in affected cells. Nature, 247, 417.
- ROWLAND, A. C., LAWSON, G. H. K. & ROBERTS, L., 1978. Intestinal adenomatosis: Histochemical and electron microscopic studies of the mucosa. Research in Veterinary Science, 24, 191-199.
- ROWLAND, A. C. & ROWNTREE, P. G. M., 1972. A haemorrhagic bowel syndrome associated with intestinal adenomatosis in the pig. The Veterinary Record, 91, 235–241.
- ROWNTREE, P. G. M., 1972. A haemorrhagic bowel syndrome in the pig. The Veterinary Record, 91, 235–241.
- SAIT, M., AITCHISON, K., WHEELHOUSE, N., WILSON, K., LAINSON, A., LONGBOTTOM, D. & SMITH, D. G. E., 2013. Genome sequence of Lawsonia intracellularis strain N343, isolated from a sow with hemorrhagic proliferative enteropathy. Genome Announcements, 1, e00027-13. doi:10.1128/genomeA.00027-13.
- SAMPIERI, F., VANNUCCI, F. A., ALLEN, A. L., PUSTERLA, N., ANTONOPOULOS, A. J., BALL, K. R., THOMPSON, J., DOWLING, P. M., HAMILTON, D. L. & GEBHART, C. J., 2013. Species-specificity of equine and porcine Lawsonia intracellularis isolates in laboratory animals. Canadian Journal of Veterinary Research, 77, 261-272.
- SAPICO, F. L., REEVES, D., WEXLER, H. M., DUNCAN, J., WILSON, K. H. & FINEGOLD, S. M., 1994. Preliminary study using species-specific oligonucleotide probe for rRNA of Bilophila wadsworthia. Journal of Clinical Microbiology, 32, 2510-2513.
- SCHMITZ-ESSER, M., HAFERKAMP, I., KNAB, S., PENZ, T., AST, M., KOHL, C., WAGNER, M. & HORN, M., 2008. Lawsonia intracellularis contains a gene encoding a functional Rickettsia-like ATP/ADP translocase for host exploitation. Journal of Bacteriology, 190, 5746-5752.
- SMITH, S. H. & MCORIST, S., 1997. Development of persistent intestinal infection and excretion of Lawsonia intracellularis by piglets. Research in Veterinary Science, 62, 6-10.
- SMITH, S. H., MCORIST, S. & GREEN, L. E., 1998. Questionnaire survey of proliferative enteropathy on British pig farms. The Veterinary Record, 142, 690–693.
- STEGE, H., JENSEN, T. K., MÖLLER, K., VESTERGAARD, K., BAEKBO, P. & JORSAL, S. E., 2004. Infection dynamics of Lawsonia intracellularis in pig herds. Veterinary Microbiology, 104, 197–206.
- STRAW, B., DEWEY, C., KOBER, J. & HENRY, S. C., 2002. Factors associated with death due to hemorrhagic bowel syndrome in two large commercial swine farms. Journal of Swine Health and Production, 10, 75-79.
- VAN DER WALT, M. L., SPENCER, B. T. & LOVEDAY, R. K., 1988. The first isolation of Campylobacter mucosalis from pigs in South Africa. Onderstepoort Journal of Veterinary Research, 55, 165–168.
- VAN DER WALT, M. L. & VAN DER LUGT, J. J., 1988. The isolation of Campylobacter hyointestinalis from pigs in South Africa. Onderspoort Journal of Veterinary Research, 55, 85-87.
- VANNUCCI, F. A., FOSTER, D. N. & GEBHART, C. J., 2012. Comparative transcriptional analysis of homologous pathogenic and non-pathogenic Lawsonia intracellularis isolates in infected porcine cells. PLoS One, 7, 1-13, e46708. doi: 10.1371/journal.pone.0046708.
- VANNUCCI, F. A., FOSTER, D. N. & GEBHART, C. J., 2013. Laser microdissection coupled with RNA-seq analysis of porcine enterocytes infected with an obligate intracellular pathogen (Lawsonia intracellularis). BMC Genomics, 14, 421. doi: 10.1186/1471-2164-14-421.
- VANNUCCI, F. A., KELLEY, M. R. & GEBHART, C. J., 2013. Comparative genome sequencing identifies a prophage-associated genomic island linked to host adaptation of Lawsonia intracellularis infections. Veterinary Research, 44, 1-9. http://www.veterinaryresearch.org/content/44/1/49.
- VANNUCCI, F. A., PUSTERLA, N., MAPES, S. M. & GEBHART, C. J., 2012. Evidence of host adaptation in Lawsonia intracellularis infections. Veterinary Research, 43, 1-9. http://veterinaryresearch.org/content/43/1/53.
- WAGNER, J. E., OWENS, D. R. & TROUTT, H. F., 1973. Proliferative ileitis of hamsters: Electron microscopy of bacteria in cells. American Journal of Veterinary Research, 34, 249–252.
- WALTER, D., GEBHART, C. J., KROLL, J., HOLCK, J. T. & CHITTICK, W., 2004. Serologic profiling and vaccination timing for Lawsonia intracellularis. Journal of Swine Health and Production, 12, 310-313.
- WATSON, E., ALBERDI, M. P., INGLIS, N. F., LAINSON, A., PORTER, M. E., MANSON, E., IMRIE, L., MCLEAN, K. & SMITH, D. G. E., 2014. Proteomic analysis of Lawsonia intracellularis reveals expression of outer membrane proteins during infection. Veterinary Microbiology, 174, 448-455.
- WATTANAPHANSAK, S., ASAWAKARN, T., GEBHART, C. J. & DEEN, J., 2008. Development and validation of an enzyme-linked immunosorbent assay for the diagnosis of porcine proliferative enteropathy. Journal of Veterinary Diagnostic Investigation, 20, 170-177.
- WATTANAPHANSAK, S., SINGER, R. S. & GEBHART, C. J., 2010. Evaluation of in vitro bactericidal activity of commercial disinfectants against Lawsonia intracellularis. Journal of Swine Health and Production, 18, 11-17.
- WEISBURG, W. G., BARNS, S. M., PELLETIER, D. A. & LANE, D. J., 1991. 16S ribosomal DNA amplification for phylogenetic study. Journal of Bacteriology, 173, 697–703.
- WILLIAMS, M. C., 1994. Faculty of Veterinary Science, University of Pretoria, Onderstepoort, South Africa.Unpublished data. In: COETZER, J. A. W., THOMSON, G. R. & TUSTIN, R. C., (eds.). Infectious Diseases of Livestock with Special Reference to Southern Africa. Oxford University Press, Southern Africa.
- WINKELMAN, N. L. & DEE, S., 1996. Ileitis: an update. Compendium of Continuing Education of Practicing Veterinarians 18, S19–S25.
- WON, G. & LEE, J., H. , 2018. Antigenic and functional profiles of a Lawsonia intracellularis protein that shows a flagellin-like trait and its immune-stimulatory assessment. Veterinary Research, 49, 1-8. https://doi.org/10.1186/s13567-018-0515-0.
- ZUMPT, I. F., 1994. Regional Veterinary Laboratory, Stellenbosch, Cape Province, South Africa.Unpublished data. In: COETZER, J. A. W., THOMSON, G. R. & TUSTIN, R. C., (eds.) Infectious Diseases of Livestock with Special Reference to Southern Africa. Cape Town: Oxford University Press, Southern Africa.




