- Infectious Diseases of Livestock
- Part 3
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Progressive atrophic rhinitis
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Progressive atrophic rhinitis
M-L PENRITH
Introduction
Progressive atrophic rhinitis (PAR) is an important disease of pigs characterized by turbinate atrophy with varying degrees of facial deformation. It is caused by infection with toxigenic strains of Pasteurella multocida. Clinical disease can cause production losses due to growth retardation in fattening pigs.17 Subclinical disease also occurs.
The aetiology of PAR was long disputed, as various causes of turbinate atrophy have been identified. Infection with Bordetella bronchiseptica can cause severe rhinitis with turbinate atrophy in piglets less than six weeks old. However, it is accepted that the progressive form of the disease, which can affect older pigs, is caused only by toxigenic strains of P. multocida,17, 22, 24, 67 although various factors including infection with B. bronchiseptica can predispose to PAR. The progressive nature of the disease probably depends upon chronic infection with toxigenic P. multocida, as turbinate atrophy experimentally induced by a single intraperitoneal administration of toxin resolved within approximately three months.57
Progressive atrophic rhinitis has probably been known since 1830, when it was described as ‘Schnüffelkrankheit’.17 It was reported from the USA in 194428 and the UK in 1954.8 It is widespread, occurring in most countries that produce pigs, including South Africa. It has recently been reported in Kenya.82 Lesions suggestive of PAR have been described in extensively kept pigs of local breed in Zimbabwe,36 although this was not confirmed by bacterial isolation.
The importance of PAR lies not only in its ability to cause production losses, but in the fact that subclinical disease is common. Herds that have never shown clinical signs of atrophic rhinitis may nevertheless harbour carriers that, introduced into herds under different management, may spread the infection. Detection of carriers is therefore of critical importance to prevent introduction into clean herds. Because even modern techniques may fail to identify infected individuals, continuous monitoring of herds is necessary to ensure freedom from PAR.
Although PAR is of importance only in pigs, toxigenic strains of P. multocida have been isolated from rabbits, humans, and calves, in which they produce similar effects of turbinate atrophy, as well as from dogs, cats, goats and poultry.44
Aetiology
A toxin produced by particular strains of P. multocida is the cause of progressive turbinate atrophy in pigs, either alone or in combination with B. bronchiseptica.17, 24, 74, 75 Pasteurella multocida is best cultured in broth media.17 Strains of both types A and D cultured from pigs, rabbits and humans have been identified that produce a thermolabile dermonecrotic protein toxin. An untypable strain was isolated from cattle with turbinate atrophy and found also to produce a dermonecrotic toxin.44 The majority of toxigenic strains isolated from pigs belong to type D, but occasionally toxigenic type A strains are reported.33, 85 Identification of the gene that encodes for the toxin indicated that it may be located in a bacteriophage, transfer of which may be able to cause non-toxigenic strains to produce toxin.7 This hypothesis was supported by the finding in another study that the toxin-encoding gene may have had a different origin from the rest of the genome of the organism studied.47
Epidemiology
Clinical PAR usually occurs in young pigs. Experimental infection has succeeded in producing typical lesions of PAR in pigs up to 16 weeks old, although infection usually occurs in the farrowing unit before piglets are four weeks old.62 Lesions decrease in severity with age, and infection of adults may result in subclinical disease with subsequent infection of piglets by their dams, 10 to 15 per cent of which may be carriers.17 The highest prevalence of carriers was found in the 6- to 12-month age group.83
Introduction into clean herds is usually by means of carriers, which may shed organisms when stressed by transport and mixing with other pigs. Molecular fingerprinting of isolates in Australia suggested that importation of infected pigs from a single source was responsible for PAR in Australia.33
Transmission is usually by direct nose-to-nose contact or aerosol, but, since toxin administered by various routes can produce lesions of PAR, and it appears that toxin can be produced in sites other than the nasal mucosa, infection by other routes may be possible.1, 2, 85 Toxigenic P. multocida is usually carried in the pharyngeal tonsils1, 2 but may be isolated from the nasal mucosa and the lungs.60 It has also been isolated from the vaginas of sows.17 Transmission between species might occur, but it has been suggested that this may not always result in disease.45 However, piglets developed turbinate atrophy after inoculation with toxigenic P. multocida cultured from a human patient.59
Although toxin produced by P. multocida is capable of producing typical lesions of PAR, colonization of the nasal mucosa is enhanced by the presence of B. bronchiseptica.17, 29, 74, 75 However, vaccination with purified P. multocida toxin alone gave good protection against severe lesions of PAR.32 Damage induced by high levels of ammonia and dust also facilitates colonization.17, 38
The severity of PAR, and indeed whether it will manifest clinically at all, depends on environmental factors and above all management. In the USA, it was demonstrated that pigs slaughtered in spring or summer (but farrowed and raised in winter) were more frequently affected with PAR.16, 77 Enclosed housing with limited, controlled ventilation was also associated with a higher incidence of PAR.16
Toxigenic P. multocida has also been associated with pneumonia in pigs.60
Pathogenesis
Progressive atrophic rhinitis results from colonization by a toxigenic strain of P. multocida, followed by elaboration of the toxin which exerts a direct effect on turbinate growth. The pharyngeal tonsil is a predilection site for colonization by P. multocida. 1, 2, 38, 72 In the absence of damage to the nasal mucosa caused by chemical effects, such as those resulting from poor air quality or infection with B. bronchiseptica, colonization by P. multocida is poor.29, 38, 66 Very high doses of P. multocida toxin as well as infection with B. bronchiseptica and various chemical mediators61 cause ciliostasis, which facilitates colonization of the nasal mucosa. Colonization of the nasal mucosa by toxigenic P. multocida results in more severe lesions,32 but is not essential for the induction of PAR: toxin produced in the tonsils or the lungs can also cause turbinate lesions.1 It has been demonstrated that not only intranasal but also subcutaneous, intramuscular and intraperitoneal administration of toxin can produce turbinate atrophy.2, 85
Colonization of the nasal mucosa by toxigenic P. multocida results in epithelial hyperplasia and atrophy of mucosal glands, accompanied by an influx of neutrophils and mononuclear cells.17 The osteoporotic changes in the turbinate bones induced by the toxin are the result of increased bone resorption and impaired osteogenesis,25, 30, 50, 66 characterized by decreased mineralization, and replacement fibrosis. Increased osteoclasia induced by P. multocida toxin has been demonstrated to result from an increase in both numbers and activity of osteoclasts.50
The increase in number is ascribed to the ability of the toxin to induce the differentiation of osteoclast- like cells from precursors in the bone marrow,51 possibly indirectly via cytokine production by a subpopulation of stromal cells.37 The marked effect on turbinate bones is probably due to the fact that these consist of endochondral bone with few trabeculae and a large amount of bone marrow that is actively haemopoietic.51 Stimulation of osteoclasia appears to be transient,25 which may explain why an increased number of osteoclasts is not always observed.
The effects on osteoblasts appear to include cytolysis and impairment of differentiation and function. There is evidence that P. multocida toxin may prevent differentiation of precursor cells into osteoblasts.63, 64, 73 Ultrastructural studies revealed degenerative changes in differentiated osteoblasts.66 In vitro studies have shown that incubation of cells with osteoblastic activity with P. multocida toxin at doses comparable to those required to induce turbinate atrophy resulted in a marked reduction in alkaline phosphatase activity,37, 78 which would account for decreased mineralization. Cytolysis induced by P. multocida toxin appears to be dose-dependent.37
The way in which P. multocida toxin causes growth retardation has not been fully eludicated. It is generally accepted that PAR can result in growth retardation.17, 57, 60 Pigs inoculated with a single dose of purified P. multocida toxin developed growth retardation within a week, which was maximal at two weeks and still observed, although reduced, at slaughter.57 The degree to which PAR results in increased time to market is not strictly correlated with the severity of lesions observed at slaughter.2, 77 It could be surmised that any condition that affects the upper respiratory tract chronically would lead to reduced feed intake and therefore poorer growth. However, experimentally induced disease appeared to have little effect on the appetite of the pigs38 and did not affect the feed conversion ratio.4 Growth hormone plasma concentrations and pituitary response have been shown to be normal in pigs suffering from natural and experimentally induced PAR,34 and the authors speculated that the toxin may exert an effect through modulation of signal transduction pathways. Adverse effects on physeal growth have been reported.3, 4 Physeal area and chondrocyte proliferation were reduced in piglets infected parenterally with P. multocida toxin.3
Clinical signs and pathology
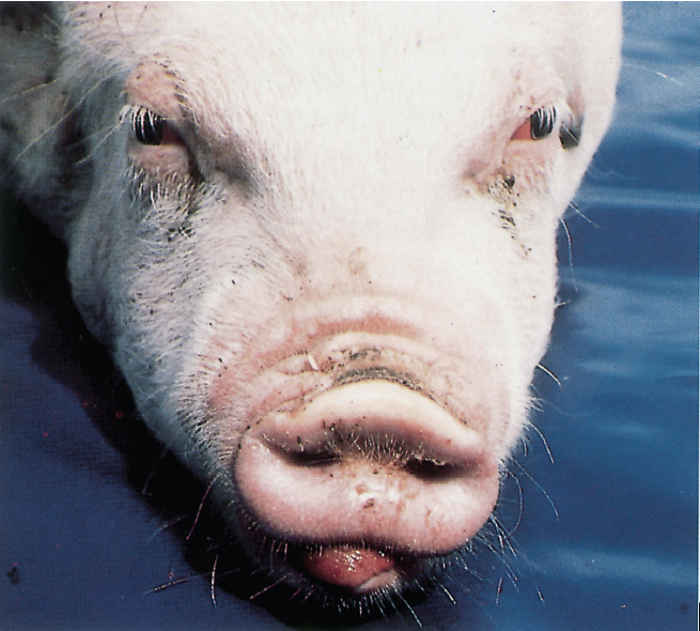
Clinical signs usually do not develop until piglets are 4 to 12 weeks of age. The first signs are sneezing and snuffling as a result of acute rhinitis. Piglets affected with PAR continue to sneeze and snuffle throughout the growing period,producing varying amounts of serous to mucopurulent nasal and ocular discharges. As a result of the ocular discharge, triangular dark stains, or tear-marks, develop below the medial canthi of the eyes. Animals that are severely affected may suffer bouts of epistaxis, which is usually unilateral, and this may be evident from the presence of blood stains on the walls of the pens. Pieces of turbinate bone may be expelled during particularly violent sneezing. As the disease progresses, deformities of the snout may become evident. Shortening of the upper jaw (brachygnathia superior) is the most commonly observed deformity, with folding of the skin over the dorsum of the snout (Figure 167.1). Turbinate lesions may be asymmetrical, with deviation of the nasal septum, which causes deviation of the snout towards the side that is more severely affected (Figure 167.2). Growth retardation and poor feed conversion may occur in severely diseased pigs.
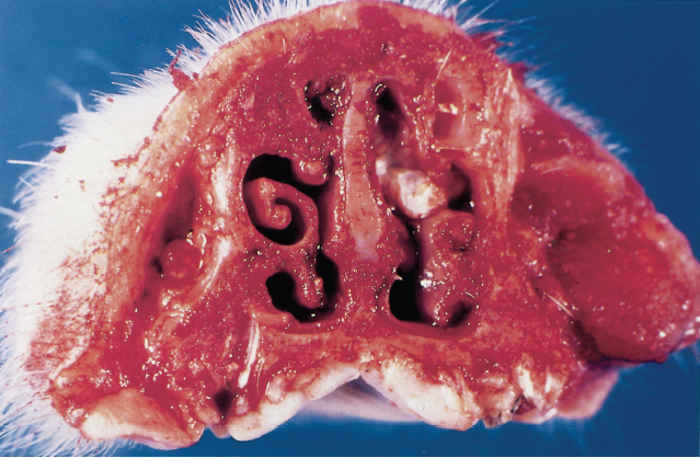
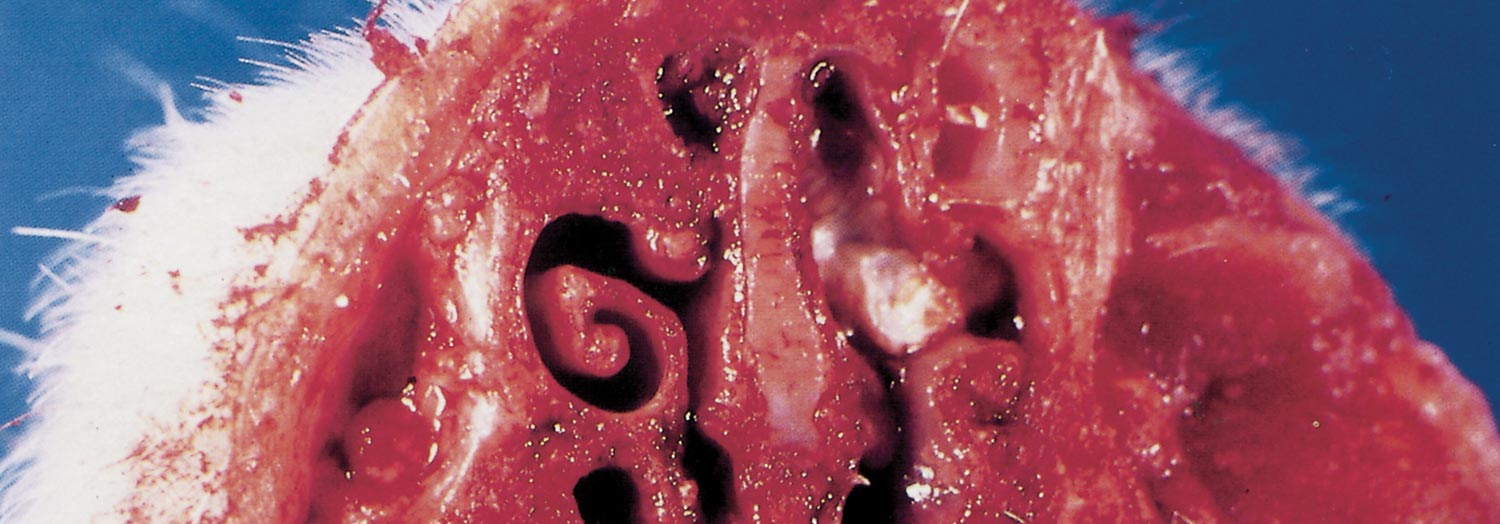
In order to evaluate the turbinate lesions, snouts of slaughtered pigs should be cut at the level of premolars 1 and 2 and examined in cross-section. Comparison with a normal snout cut at the same level facilitates evaluation of the severity of the lesions.
The ventral conchae are the first to be affected, and may be so reduced in size to be absent. In severe cases the dorsal and ventral conchae are markedly reduced, and in very severely affected pigs the conchae may be absent. The lesions exhibit varying degrees of asymmetry. The nasal septum may be normal or deflected to one side, depending on the degree of asymmetry of the lesions.
Other organs are usually normal, but signs of pneumonia may be present, as infection with Mycoplasma hyopneumoniae has been reported to predispose to PAR, and toxigenic P. multocida has also been isolated from pneumonic lesions. 60 The toxin of P. multocida has also been demonstrated to cause thickening of the epithelium of the urinary bladder.39 Experimental infection with high doses of purified toxin resulted in icterus due to severe hepatocellular degeneration and necrosis, and gastric ulceration and polioencephalomalacia were also observed.27
Diagnosis
Pigs suffering from clinical PAR may be smaller than their litter mates, have a dull hairy coat, and exhibit varying degrees of facial deformation as described above. Tear stains are often present and are suggestive of but not pathognomonic for PAR.
Because the disease may be subclinically present in herds that have never shown any clinical signs of PAR, abattoir monitoring is frequently used to detect lesions suggestive of PAR. To facilitate diagnosis, various methods of scoring snouts have been developed, ranging from subjective scoring severity lesions on a 0 to 5 scale to more sophisticated methods to try to achieve an objective evaluation. The latter include morphometric approaches10, 26, 52 and computerized tomography scans.41 However, the cut-off point at which the lesion is classified as PAR remains a problem.35 In order to make a definitive diagnosis, involvement of toxigenic P. multocida must be confirmed in the laboratory.
The highest number of organisms usually occurs in the tonsils,18, 38, 72 making this the specimen of choice for isolation and identification of the organism.
Traditionally, confirmation of the diagnosis depended upon isolation of the organism, and subsequent testing in guinea pigs or mice to establish the production of dermonecrotic toxin, but this method has been superseded by an in vitro test in which Vero cells are employed to avoid the use of guinea pigs.70 These methods are slow, laborious and expensive. Culture of P. multocida is hampered by difficulty of growing it on solid media.17 The use of liquid or selective media is recommended.17, 46 More modern techniques, in particular the polymerase chain reaction (PCR) technique are currently used.43, 55 An ELISA has been developed in which monoclonal antibodies are used to detect the toxin.14, 53, 54
Both the ELISA and the PCR are highly sensitive and specific,6 as well as being faster than conventional tests. The results of a study on the age distribution of pigs found to be positive with the antigen-detecting ELISA suggested that pigs 6 to 12 months of age should be used in herd screening tests, as many pigs younger than this were negative, and the number of positive pigs also declined with age.83
Since specific antibodies against P. multocida toxin are produced, tests to determine serum antibody levels can detect unvaccinated herds that have been exposed to infection. Serum neutralization tests and an ELISA have been developed.5, 11, 31 A skin test based on the presence of antitoxin antibodies has been developed.15 These tests are more useful for herd screening than for the detection of individual carrier pigs, since they depend on the response of the pig to antigenic stimulation, and furthermore do not necessarily imply active infection.
In the absence of any clinical signs of PAR, detection of toxigenic P. multocida does not constitute a definitive diagnosis of PAR. It has been suggested, on the basis of identification of toxigenic P. multocida in the prolonged absence of disease in a closed herd in Ireland, that the strain identified, which differed from the usual strain that causes PAR in the UK, may have been derived from a different host and therefore did not spread and cause disease in pigs.45
Differential diagnosis
Acute rhinitis in young piglets has various causes, including B. bronchiseptica, inclusion body rhinitis caused by porcine cytomegalovirus, dusty feed, or poor environmental conditions. The diagnosis of PAR depends on isolation or detection of toxigenic P. multocida, and progression to development of lesions typical of PAR. Later stages of the disease must be differentiated from other causes of facial deformation such as abscesses, trauma to the nasal bones, and congenital conditions. These conditions are likely to affect individual animals only.
Control
Treatment during an outbreak of atrophic rhinitis is aimed at controlling the infection in pigs with clinical signs and preventing its spread in the herd by reducing the overall bacterial load.17 This should be accompanied by preventive measures that usually include vaccination and medication of sows, and improving the environment for the pigs, with particular attention to management of ventilation and housing.17
Various vaccines provide protection. Combinations of killed B. bronchiseptica cells and inactivated P. multocida organisms and toxin have been used with good effect.23, 42, 48, 76, 81 The P. multocida type D toxoid cross-protects against type A.76 Good protection with this type of vaccine was reported after sows were vaccinated five to six and two to three weeks before farrowing,79 but a single vaccination in weaned piglets did not protect against challenge with cellfree toxin. The use of different adjuvants does not appear to affect vaccine efficacy.42, 65 Toxin purified with monoclonal antibodies alone is capable of protecting piglets against infection with B. bronchiseptica and toxigenic P. multocida.31, 80 Promising results have been obtained with a recombinant vaccine.13, 58, 71 Commercial vaccines are widely available. In South Africa they are often used in conjunction with medication of sows with oxytetracycline.
Medication may be used alone or in combination with vaccination to treat or prevent PAR. A wide variety of antimicrobials has proved effective against PAR. Potentiated sulphonamides, oxytetracycline, penicillin/streptomycin combinations, lincomycin/spectinamycin combinations, tilmicosin, and enrofloxacin have all been reported to be efficacious.9, 12, 17, 40, 69
Various régimes have been advocated, including in-feed or parenteral medication of sows during the last month of pregnancy, and parenteral, intranasal or in-feed medication of piglets and growers.9, 12, 40, 69 Combining vaccination of sows with antimicrobial treatment has been reported to give the best results.49, 68 When toxigenic P. multocida is isolated during an outbreak, the performance of an antibiogram is advisable in order to ensure that an antimicrobial of optimal sensitivity is selected. If an antibiogram is not carried out, a broad-spectrum antibiotic should be selected according to affordability and practicality, and administered according to the manufacturer’s instructions.
To eradicate PAR from infected sow herds, vaccination over a period of at least four years combined with testing nasal and tonsillar swabs, using polymerase chain reaction (PCR) or a combination of the PCR and antigen-detecting ELISA, has been advocated.19, 21 This should be combined with purchasing new stock only from herds known to be free of PAR. Screening to ensure freedom from carriers is performed by obtaining nasal and tonsillar swabs, as well as tonsillar biopsies, from the entire breeding herd20 to ensure freedom from infection.
Measures are necessary to prevent introduction of PAR into naive herds by carrier pigs. New stock should be acquired from herds known to be free of the disease, if an accreditation system exists.
New pigs should be quarantined separately from the herd for at least six weeks, and nasal and tonsillar swabs taken for bacteriological testing should prove negative before the pigs are introduced into the herd.
Management is very important in the prevention of clinical PAR. Poor air quality in the farrowing unit, mixing of animals of different ages, and high stocking densities, particularly in continuous flow systems, predispose to PAR.56, 84 Reducing the level of toxigenic P. multocida in the farrowing unit and optimizing piglet defence mechanisms are helpful in controlling PAR.62 Ensuring that the environmental temperature is correct and excluding factors such as draughts that may stress piglets and lower their resistance contribute to its control.17 It is possible that artificial insemination could contribute to the control of PAR by eliminating contact with carrier boars.17 Modified medicated early weaning, all-in-all-out systems, strict hygienic measures, and improvement of the environment to ensure that ammonia levels are acceptable and that excessive dust is not present in the atmosphere or generated by the feed, are helpful in the control of PAR.
References
- ACKERMANN, M.R., CHEVILLE, N.F. & GALLAGHER, J.E., 1991. Colonisation of the pharyngeal tonsil and respiratory tract of the gnotobiotic pig by a toxigenic strain of Pasteurella multocida type D. Veterinary Pathology, 28: 267–274.
- ACKERMANN, M.R., DEBEY, M.C., REGISTER, K.B., LARSON, D.J. & KINYON, J.M., 1994. Tonsil and turbinate colonization by toxigenic and nontoxigenic strains of Pasteurella multocida in conventionally-raised swine. Journal of Diagnostic Investigation, 6, 375–377.
- ACKERMANN, M.R., REGISTER, K.B., STABEL, J.R., GWALTNEY, S.M., HOWE, T.S. & RIMLER, R.B., 1996. Effect of Pasteurella multocida toxin on physeal growth in young pigs. American Journal of Veterinary Research, 57, 848–852.
- ACKERMANN, M.R., STABEL, J.R., PETTIT, R.K., JACOBSON, C.D., ELMQUIST, J.K., REGISTER, K.B., RIMLER, R.B. & HILTON, J.A., 1995. Reduced physeal area and chondrocyte proliferation in Pasteurella multocida toxin-treated rats. Veterinary Pathology, 32, 674–682.
- ALT, M., BECHMANN, G. & SCHÖSS, P., 1992. Comparison of EBL cells and ELISA in the serological diagnosis of progressive atrophic rhinitis (PAR). Proceedings of the 12th IPVS Congress, The Hague, The Netherlands, 17–20 August 1992, 12, 153.
- AMIGOT, J.A., TORREMORELL, M. & PIJOAN, C., 1998. Evaluation of techniques for the detection of toxigenic Pasteurella multocida strains from pigs. Journal of Veterinary Diagnostic Investigation, 10, 169–173.
- ANDRESEN, L.O., PETERSEN, S.K., CHRISTIANSEN, C. & NIELSEN, J.P., 1990. Studies on the location of the Pasteurella multocida toxin gene, toxA. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 60.
- ANON, 1954. New pig disease in Britain. The Veterinary Record, 66, 316.
- BÄCKSTRÖM, L., CHUNG, W.B., COLLINS, M.T. & OSE, E., 1992. Efficacy of tilmicosin for control of atrophic rhinitis. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands, 17–20 August 1992, 12, 161.
- BARFOD, K., SORENSON, V. & NIELSEN, J.P., 1990. Methods of evaluation of the degree of atrophic rhinitis. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 70.
- BECHMANN, G. & SCHÖMSS, P., 1990. Neutralizing activity against Pasteurella multocida toxin in sera of pigs with atrophic rhinitis. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 50.
- BINDER, S., LACKHOFF, A., LE, N.B., BERNER, H. & BAUER, J., 1992. Efficacy of food-medication with tilmicosin in respiratory disorders of swine. Proceedings of the 12th IPVS Congress,The Hague, The Netherlands, 17–20 August 1992, 12, 162.
- BORDING, A., NYMARK, K., SMIDT, E. & HJALAGER, B., 1992. Field studies with a new, genetically engineered vaccine against progressive atrophic rhinitis. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands, 17–20 August 1992, 12, 179.
- BOWERSOCK, T.L., HOOPER, T. & POTTENGER, R., 1992. Use of ELISA to detect toxigenic Pasteurella multocida in atrophic rhinitis in swine. Journal of Veterinary Diagnostic Investigation, 4, 419–422.
- BREUER, J. & SCHIMMELPFENNIG, H., 1990. Skin-testing for P. multocida antitoxic antibodies in breeding herds. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 51.
- COWART, R.P., BOESSEN, C.R. & KLIEBENSTEIN, J.B., 1992. Patterns associated with season and facilities for atrophic rhinitis and pneumonia in slaughtered swine. Journal of the American Veterinary Medical Association, 200, 190–193.
- DE JONG, M.F. 1992. (Progressive) atrophic rhinitis. In: leman, a.d., straw, b., mengeling, w.l., d’allaire, s. & taylor, d.j., (eds). Diseases of Swine. 7th edn. Ames: Iowa State University Press.
- DE JONG, M.F. & BRAAMSKAMP, J., 1994. Atrophic rhinitis herd monitoring: The investigation of toxinogenic Pasteurella multocida from the tonsils of culled sows collected in the slaughterhouse. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 161.
- DE JONG, M.F., HUNNEMAN, W.A., MEKKES, D. & BANNISETH, T.V., 2000. Eradication of atrophic rhinitis toxigenic Pasteurella multocida in ART vaccinating sow herds with a PCR test. Proceedings of the 16th IPVS Congress, Melbourne, Australia, 17–20 September 2000, 477.
- DE JONG, M.F., KAMP, E. & BOKKEN, G., 1994. Selecting sows harbouring AR toxigenic Pasteurella multocida by a PCR-test to eliminate progressive AR in a breeding herd. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 167.
- de jong, m.f., kamp, e., van der schoot, a. & von banniseth, t., 1996. Elimination of AR toxigenic Pasteurella from infected sow herds by a combination of ART vaccination and testing sows with a PCR and ELISA test. Proceedings of the 14th IPVS Congress, Bologna, Italy, 7–10 July 1996, 245.
- DE JONG, M.F. & NIELSEN, J.P., 1990. Definition of progressive atrophic rhinitis. The Veterinary Record, 27, 93.
- DESCAMPS, J., CHARLIER, P., JOLIE, R. & DE ROOSE, P., 1990. Efficacy of a vaccine against atrophic rhinitis under field conditions. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 69.
- DOMINICK, M.A. & RIMLER, R.B., 1986. Turbinate atrophy in gnotobiotic pigs intranasally inoculated with protein toxin isolated from type D Pasteurella multocida. American Journal of Veterinary Research, 47, 1532–1536.
- DOMINICK, M.A. & RIMLER, R.B., 1988. Turbinate osteoporosis in pigs following intranasal inoculation of purified Pasteurella multocida toxin: Histomorphometric and ultrastructural studies. Veterinary Pathology, 25, 17–27.
- DONE, J.T., UPCOTT, D.H., FREWIN, D.C. & HERBERT, C.N., 1984. Atrophic rhinitis: Snout morphometry for quantitative assessment of conchal atrophy. The Veterinary Record, 113, 33–45.
- DOSTER, A.R., FRANTZ, J.C., BROWN, A.L., HUSEMAN, B.R. & HOGG, A., 1990. Effects of Pasteurella multocida serotype D dermonecrotic toxin in swine. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 72.
- DOYLE, L.P., DONHAM, C.R. & HUTCHINGS, L.M., 1944. Report of a type of rhinitis in swine. Journal of the American Veterinary Medical Association, 105, 132.
- ÉLIÁS, B., ALBERT, M., TUBOLY, S. & RAFAI, P., 1992. Interaction between Bordetella bronchiseptica and toxigenic Pasteurella multocida on the nasal mucosa of SPF piglets. Journal of Veterinary Medical Science, 54, 1105–1110.
- FELIX, R., FLEISCH, H. & FRANDSEN, P.L., 1992. Effect of Pasteurella multocida toxin on bone resorption in vitro. Infection and Immunity, 60, 4984–4988.
- FOGED, N.T., NIELSEN, J.P., & BARFOD, K., 1990. The use of ELISAdetermination of Pasteurella multocida toxin antibodies in the control of progressive atrophic rhinitis. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 49.
- FOGED, N.T., NIELSEN, J.P. & JORSAL, S.E., 1989. Protection against progressive atrophic rhinitis by vaccination with Pasteurella multocida toxin purified by monoclonal antibodies. The Veterinary Record, 125, 7–11.
- GARDNER, I.A., KASTEN, R., EAMENS, G.J., SNIPES, K.P. & ANDERSON, R.J., 1994. Molecular fingerprinting of Pasteurella multocida associated with atrophic rhinitis in swine herds. Journal of Veterinary Diagnostic Investigation, 6, 442–447.
- GHOSHAL, N.G., NIYO, Y. & TRENKLE, A.H., 1991. Growth hormone concentrations in plasma of healthy pigs and pigs with atrophic rhinitis. American Journal of Veterinary Research, 52, 1684–1687.
- GOODWIN, R.F.W., 1988. Monitoring for atrophic rhinitis: The problem of higher snout scores. The Veterinary Record, 123, 566–568.
- GUPTA, S.K., RUZIWA, S.D., ERLWANGER, K.H. & DZAMA, K., 1999. Atrophic rhinitis in an indigenous Zimbabwe pig (mukota)—a case report. Proceedings of the IXth International Conference of Association of Institutions of Tropical Veterinary Medicine, Harare, Zimbabwe, 14–18 September 1998, 2, 611.
- GWALTNEY, S.M., GALVIN, R.J.S, REGISTER, K.B., RIMLER, R.B. & ACKERMANN, M.R., 1997. Effects of Pasteurella multocida toxin on porcine bone marrow cell differentiation into osteoclasts and osteoblasts. Veterinary Pathology, 34, 421–430.
- HAMILTON, T.D.C, ROE, J.M. & WEBSTER, A.J.F., 1998. The synergistic role of gaseous ammonia in the etiology of Pasteurella multocida-induced atrophic rhinitis in pigs. Proceedings of the 15th IPVS Congress, Birmingham, England, 5–9 July 1998, 2, 144.
- HOSKINS, I.C., THOMAS, L.H. & LAX, A.J., 1997. Nasal infection with Pasteurella multocida causes proliferation of bladder epithelium in gnotobiotic pigs. The Veterinary Record, 140, 22.
- IKOMA, H., 1994. Control of atrophic rhinitis with enrofloxacin. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 170.
- JOLIE, R. & THACKER, B., 1990. Comparison of atrophic rhinitis morphometric measurements and macroscopic grades of nasal cross sections on computerized tomography scans in pigs. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July1990, 2, 53.
- KABAY, M.J., MERCY, A.R., LLOYD, J.M. & ROBERTSON, G.M., 1992. Vaccine efficacy for reducing turbinate atrophy and improving growth rate in piggeries with endemic atrophic rhinitis. Australian Veterinary Journal, 69, 101–103.
- KAMP, E.M., BOKKEN, G.C.A.M., VERMEULEN, T.M.M., DE JONG, M.F., BUYS, H.E.C.M., REEK, F.H. & SMITS, M.A., 1996. A specific and sensitive PCR assay suitable for large-scale detection of toxigenic Pasteurella multocida in nasal and tonsillar swabs specimens of pigs.Journal of Veterinary Diagnostic Investigation, 8, 304–309.
- KAMP, E.M., TER LAAK, E.A. & DE JONG, M.F., 1990. Atypical pasteurella strains producing a toxin similar to the dermonecrotic toxin of Pasteurella multocida subspecies multocida. The Veterinary Record, 126, 434–437.
- KAVANAGH, N.T., 1994. Isolation of toxigenic Pasteurella multocida type D from pigs in a herd free from progressive atrophic rhinitis in Ireland. The Veterinary Record, 134, 218–219.
- LARIVIERE, S., LEBLANC, L., MITTAL, K.R. & MARTINEAU, G.-P., 1993. Comparison of isolation methods for the recovery of Bordetella bronchiseptica and Pasteurella multocida from the nasal cavities of piglets. Journal of Clinical Microbiology, 31, 364–367.
- LAX, A.J. & CHANTER, N., 1990. Cloning of the toxin gene from Pasteurella multocida and its role in atrophic rhinitis. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 61.
- MAGYAR, T., KOVACS, F. & VESTERGAARD-NIELSEN, K., 2000. Vaccine efficacy comparison against combined infections with Bordetella bronchiseptica and Pasteurella multocida in newborn piglets. Proceedings of the 16th IPVS Congress, Melbourne, Australia, 17–20 September 2000, 479.
- MARTELLI, P., SALA, V. & POZZI, P., 1994. Evaluation of a protocol to control a progressive atrophic rhinitis outbreak. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 171.
- MARTINEAU-DOIZÉ, B., CAYA, I., GAGNÉ, S., JUTRAS, I. & DUMAS, G., 1993. Effects of Pasteurella multocida toxin on the osteoclast population of the rat. Journal of Comparative Pathology, 108, 81–91.
- MARTINEAU-DOIZÉ, B. & JUTRAS, I., 1994. Effects of Pasteurella multocida toxin on osteoclast formation in long-term culture of bone marrow cells. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 164.
- MARTINEAU-DOIZE, B., LAROCHELLE, R., BOUTIN, J. & MARTINEAU, G.-P., 1990. Atrophic rhinitis caused by toxigenic Pasteurella multocida type D: Morphometric analysis. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland,1–5 July 1990, 2, 63
- MATTSSON, S., FOGED, N.T. & SÖDERLIND, O., 1992. Evaluation of the 1-step DAKO PMT ELISA for detection of toxin-producing Pasteurella multocida. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands,17–20 August 1992, 12, 173.
- MATTSSON, S. & SÖDERLIND, O., 1990. Detection of Pasteurella multocida strains in nasal swabs from pigs using an antitoxin ELISA. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland,1–5 July 1990 2, 79.
- NAGAI, S., SOMENO, S. & YAGIHASHI, T, 1994. Differentiation of toxigenic from nontoxigenic isolates of Pasteurella multocida by polymerase chain reaction. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 163.
- NIELSEN, J.P., 1990. The influence of herd epidemiological factors on atrophic rhinitis in pig herds in Denmark. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 64.
- NIELSEN, J.P. & FOGED, N.T., 1992. Duration of turbinate atrophy and growth retardation in piglets challenged with Pasteurella multocida toxin. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands, 17–20 August 1992, 12, 154.
- NIELSEN, J.P., FOGED, N.T., SORENSEN, V., BARFOD, K., JENSEN, A.B. & PETERSEN, S.K., 1990. Protection against progressive atrophic rhinitis with a recombinant Pasteurella multocida derivative. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 55.
- NIELSEN, J.P. & FREDERIKSEN, W., 1990. Atrophic rhinitis in pigs caused by a human isolate of toxigenic Pasteurella multocida. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 75.
- NIELSEN, J.P., KOLKMAN, P. & ROSENDAL, S., 1992. Infection with toxigenic Pasteurella multocida and growth rate in a cohort of pigs. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands, 17–20 August 1992, 12, 156.
- NIELSEN, J.P. & ROSENDAL, S., 1994. Ciliostasis in the pathogenesis of progressive atrophic rhinitis. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 119.
- NIELSEN, U., 1990. The influence of herd epidemiological factors on atrophic rhinitis in pig herds in Denmark. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland,1–5 July 1990, 2, 64.
- OHNISHI, T., HORIGUCHI, Y., MASUDA, M., SUGIMOTO, N. & MATSUDA, M., 1998. Pasteurella multocida toxin and Bordetella bronchiseptica dermonecrotizing toxin elicit similar effects on cultured cells by different mechanisms. Journal of Veterinary Medical Science, 60, 301–305.
- ONO, E., KOIMARU, H., NAKAI, T. & KUME, K., 1992. Effects of dermonecrotic toxin from Bordetella bronchiseptica or Pasteurella multocida on ossteoblastic line cells. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands, 17–20 August 1992, 12, 170.
- OSTLE, A.G., COYLE, D., FRANK, C., KREGNESS, B., REHDER, J. & WELTER, M., 1994. Comparative evaluation of two commercial atrophic rhinitis vaccines. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 169.
- PEDERSEN, K.B. & ELLING, F., 1984. The pathogenesis of atrophic rhinitis in pigs induced by toxigenic Pasteurella multocida. Journal of Comparative Pathology, 94, 203–214.
- PEDERSEN, K.B., NIELSEN, J.P., FOGED, N.T., ELLING, F., NIELSEN, N.C. & WILLEBERG, P., 1988. Atrophic rhinitis in pigs: Proposal for a revised definition. The Veterinary Record, 122, 190–191.
- PEJSAK, Z., HOGG, A., FOREMAN, K. & WASINSKA, B., 1990. The effect of terramycin/LA in combination with a Bordetella/Pasteurella vaccine in controlling atrophic rhinitis in swine. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland,1–5 July 1990, 2, 76.
- PEJSAK, Z., WASINSKA, B., LIPOWSKI, A. & SADOCH, L., 1990. An evaluation of some selected chemotherapeutics in the treatment of atrophic rhinitis. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland, 1–5 July 1990, 2, 76.
- PENNINGS, A.M.M.A. & STORM, P.K., 1984. A test in Vero cell monolayers for toxin production by strains of Pasteurella multocida isolated from pigs suspected of having atrophic rhinitis. Veterinary Microbiology, 9, 503–508.
- PETERSEN, S., BORDING, A., FOGED, N., FRANDSEN, P.L., NIELSEN, J.P. & SORENSON, V., 1990. Development of a recombinant vaccine against progressive atrophic rhinitis in pigs. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland,1–5 July 1990, 2, 54.
- PIFFER, I.A., CASTRO, A.F.P. & BRITO, J.R.F., 1992. Association of toxigenic type D P. multocida (Pm) to the upper respiratory tract of swine. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands, 17–20 August 1992, 12, 172.
- ROZENGURT, E., HIGGINS, T., CHANTER, N., LAX, A.J. & STADDON, J.M., 1990. ,em>Pasteurella multocida toxin is a novel potent mitogen for cultured fibroblasts. Proceedings of the 11th IPVS Congress, Lausanne, Switzerland,1–5 July 1990, 2, 66.
- RUTTER, J.M., 1983. Virulence of Pasteurella multocida in atrophic rhinitis of gnotobiotic pigs infected with Bordetella bronchiseptica. Research in Veterinary Science, 34, 287–295.
- RUTTER, J.M. & ROJAS, X., 1982. Atrophic rhinitis in gnotobiotic piglets: Differences in the pathogenicity of Pasteurella multocida in combined infections with Bordetella bronchiseptica. The Veterinary Record, 110, 531–535.
- SAKANO, T., OKADA, M., TANEDA, A., MUKAI, T. & SATO, S., 1997. Effect of Bordetella bronchiseptica and serotype D Pasteurella multocida bacterin-toxoid on the occurrence of atrophic rhinitis after experimental infection with B. bronchiseptica and toxigenic type A P. multocida. Journal of Veterinary Medical Science, 59, 55–57.
- SCHEIDT, A.B., MAYROSE, V.B., HILL, M.A., CLARK, L.K., EINSTEIN, M.E., FRANTZ, S.F., RUNNELS, L.J. & KNOX, K.E., 1992. Relationship to growth performance of pneumonia and atrophic rhinitis lesions detected in pigs at slaughter. Journal of the American Veterinary Medical Association, 200, 1492–1496.
- STERNER-KOCK, A., LANSKE, B., ÜBERSCHÄR, S. & ATKINSON, M.J., 1995. Effects of the Pasteurella multocida toxin on osteoblastic cells in vitro. Veterinary Pathology, 32, 274–279.
- TRIGO, E., REESE, L. & SIMONSON, R., 1992. Protection of piglets against atrophic rhinitis induced by the toxin of Pasteurella multocida. Proceedings of the 12th IPVS Congress, The Hague, The Netherlands,17– 20 August 1992, 12, 164.
- VOETS, M., KERSTEN, A., DE REE, J.M., CRAUWELS, P. & STUURMAN, D., 1994. Efficacy and safety of an experimental A.R.-vaccine under controlled conditions. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 124.
- VOETS, M.T., KLAASSEN, C.H.L., CHARLIER, P., WISEMAN, A. & DESCAMPS, J., 1992. Evaluation of an atrophic rhinitis vaccine under controlled conditions. The Veterinary Record, 130, 549–553.
- WABACHA, J.K., MARIBEI, J.M., THAIYA, A.G., MUNYUA, S.J.M., KARANJA, D.N. & NJOROGE, E.M., 2000. Progressive atrophic rhinitis in a medium-scale pig farm in Kiambu, Kenya. Journal of the South African Veterinary Association, 71, 122–124.
- WALLGREN, P., MATTSSON, S., RABE, J., LINDBLAD, M., MOLANDER, B. & WIERUP, M., 1994. Age distribution of pigs carrying toxin-producing Pasteurella multocida in herds affected with atrophic rhinitis. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 122.
- WALLGREN, P., MATTSSON, S., STAMPE, M., MOLANDER, B., LINDBLAD, M. & WIERUP, M., 1994. Control of infections with toxin-producing Pasteurella multocida in herds affected with atrophic rhinitis. Proceedings of the 13th IPVS Congress, Bangkok, Thailand, 26–30 June 1994, 122.
- WU, C.-C., LIU, C.-I., HSU, T.-H. & SHEN, J.-H., 1992. The experimental study on atrophic rhinitis in pigs induced by a toxin related with Pasteurella multocida type D. Proceedings of the 12th IPVS Congress,The Hague, The Netherlands, 17–20 August 1992, 12, 171.




