- Infectious Diseases of Livestock
- Part 3
- Escherichia coli infections
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Escherichia coli infections
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Escherichia coli infections
C L GYLES AND M M HENTON
Introduction
Escherichia coli predominates among the facultatively anaerobic bacteria in the intestine of many animal species. 168 Most E. coli strains are nonpathogenic but several subsets of the species have become pathogenic through the acquisition of genes that encode virulence factors, many of which are encoded on extrachromosomal genetic elements, temperate bacteriophages, or large transmissible blocks of genes called pathogenicity islands.37, 61, 73, 74, 111, 114, 115 Escherichia coli infections in livestock may be enteric or extraintestinal. The enteric infections involve colonization of the intestine and the action of toxins and/or signal transduction events. Enteric colibacillosis occurs most commonly in new-born animals in intensive farming systems, but only rarely in animals raised under extensive conditions. The most common clinical manifestation of the enteric infections is diarrhoea. Extraintestinal infections include septicaemia, oedema disease, mastitis, uterine infection, and abortion.
Escherichia coli infections occur worldwide,2, 23, 69, 75, 85, 92, 187, 189, 191 and can be of major economic significance. In southern Africa, enteric colibacillosis occurs most commonly in piglets, and less frequently in calves, lambs and kids.75, 76
Aetiology
Escherichia coli is a Gram-negative, straight, usually motile rod 1,1 to 1,5μm wide and 2 to 6 μm long, that belongs to the Enterobacteriaceae.16, 32, 145 It is facultatively anaerobic, having both respiratory and fermentative types of metabolism, and some strains possess peritrichous flagellae.
The organisms form large, pink colonies on MacConkey agar and are typically indole positive, and urease, citrate and hydrogen sulfide negative.16 Most colonies on nutrient agar are either smooth or rough, but intermediate forms and mucoid colonies do occur. Smooth forms are low-convex and shiny, possess an entire, regular circumference, and are easily dispersible in saline, while rough colonies are dry, have a dull appearance, are usually flattish, have an irregular outline, and are difficult to disperse in saline.
As a result of mutation, rough forms have lost the polysaccharide side chains of the lipopolysaccharide of the outer membrane of the cell wall. They autoagglutinate in saline and are therefore impossible to serotype by conventional means.
Escherichia coli is distinguishable from Citrobacter in that citrate cannot be used as its sole carbon source. Lactose is fermented by most strains of E. coli, and the colonies are therefore pink on MacConkey agar. Escherichia coli is usually a strong indole producer, which distinguishes it from Enterobacter. Unlike Klebsiella, E. coli does not ferment inositol or adonitol.16
An international serotyping scheme has been developed on the basis of O (the polysaccharide portion of the cell wall lipopolysaccharide), K (capsular polysaccharide), and H (flagellar protein) antigens.108, 146 The O antigens have been assigned numbers from 1 to 173, the K antigens 1 to 103, and the H antigens 1 to 50. If only the ‘O’ antigen of an isolate is identified it is said to belong to a particular ‘O’ serogroup. Isolates with the same serotype usually have many features in common but may have important differences as well. Protein appendages called pili or fimbriae constitute a fourth type of surface antigen which is often used in characterization of pathogenic E. coli.8, 37, 57, 105, 125, 131 Many of these are critical virulence factors which allow the E. coli to bind to specific pilus receptors on the surface of host epithelial cells in the intestine or urinary bladder. The fimbrial antigens have ‘F’ designations but other names which preceded the F designation are sometimes used.
Because E. coli are normal enteric flora and also enteric pathogens of livestock, steps need to be taken to assess whether an isolate from the faeces or intestine of an animal with diarrhoea or dysentery is a pathogen. Serotype is often a strong indicator of pathogenicity,72, 108, 146 but serotyping can be done in only a limited number of laboratories that have the appropriate complete collection of specific absorbed antisera and the relevant control strains. The best indicator of pathogenicicty is the possession of virulence factors such as enterotoxins, cytotoxins, colonization pili, and adhesins.1, 7, 8, 19, 37, 39, 57, 71, 73, 131, 136, 164
The term pathotype is used to describe a group of E. coli with the same array of virulence factors. There may be different pathotypes within a serotype or a single pathotype in several serotypes. Other subtyping schemes that may be used for comparing E. coli isolates include biotyping, phage typing, plasmid profiling, and pulsed field gel electrophoresis.16, 77, 103, 108
The types of E. coli that are responsible for enteric diseases in livestock are enterotoxigenic E. coli (ETEC), enteropathogenic E. coli (EPEC), and Shiga toxin-producing E. coli (STEC) (Table 150.1).106, 108, 137 An additional category of E. coli not listed in Table 150.1 and suggested to be diarrhoeagenic in cattle is a group of E. coli which produce cytotoxic necrotizing factor (CNF) and are referred to as necrotizing E. coli (NTEC).39, 40
Enterotoxigenic E. coli (ETEC)
Escherichia coli in this category produce one or more enterotoxins and colonize the small intestine in a species-specific manner by means of pili. Enterotoxigenic E. coli are the most frequent cause of enteric colibacillosis in calves and pigs.64, 135, 183, 184
Most bovine ETEC are non-haemolytic, mucoid and K99-positive and produce heat stable enterotoxin STa.2, 72 The major O groups of bovine ETEC that have been identified worldwide are O8, O9, O20, O64 and O101. The O9 and O101 strains produce not only K99 (F5) pili but also F41 pili.37 In South Africa, strains of O9:K28 and O9:K30 usually occur in calves under one week of age, with the majority being less than three days old. Strains of O78:K80, O26:K60 and O141:K85 occur in neonatal animals and also in calves up to one month old, and in aborted foetuses.
Enteropathogenic E. coli (EPEC)
Enteropathogenic E. coli are nontoxigenic E. coli which induce a characteristic attaching and effacing lesion in the small and large intestines, and adhere to HEp-2 cells (human laryngeal epithelial) and HeLa (human cervical carcinoma) cells in a pattern called localized adherence.137, 158
They have been been implicated in diarrhoea in humans, calves, lambs, rabbits and pigs,25, 92, 93, 117, 137, 151, 191 and typically possess a large virulence plasmid called the enteropathogenic E. coli adherence factor, which encodes bundle-forming pili.
Shiga toxin-producing E. coli (STEC)
Shiga toxin-producing E. coli are bacteria that produce at least one cytotoxic protein called Shiga toxin (Stx), Shiga-like toxin or verotoxin.99, 137, 142 They are implicated in enteric and systemic diseases in humans, in oedema disease in pigs, and in diarrhoea and dysentery in calves and goats.10, 26, 63, 92, 93, 137, 181 Those STEC which are implicated in severe disease in humans are called enterohaemorrhagic E. coli (EHEC), because of their ability to cause haemorrhagic lesions in the large intestine.98, 106, 137 STEC in cattle are more important as causes of human disease than they are as causes of disease in cattle.21, 137, 182 In many countries E. coli serotype O157:H7 is the major STEC implicated in human disease. This serotype is common in the intestine of cattle and can be used to induce experimental disease, but is not associated with naturally occurring disease in cattle.35
Septicaemic E. coli
Escherichia coli that cause septicaemia are not as well characterized as those implicated in enteric diseases. The most common characteristics are their ability to resist killing by serum,resistance to phagocytosis, and ability to obtain iron from low iron environments.46, 47, 78, 107, 138, 139, 180
Table 150.1 Characteristics of types of E. coli that cause diarrhoea in humans
| TYPE OF E. COLI | CHARACTERISTICS |
|---|---|
| Enterotoxigenic E. coli (ETEC) | Produce at least one enterotoxin (STa, STb, LT or combinations) Colonize by pili Do not cause structural damage to the intestine |
| Enteropathogenic E. coli (EPEC) | Induce attaching/effacing (AE) lesions in the intestine Produce no classical protein toxin Possess a 60 Mda plasmid called EAF |
| Shiga toxin-producing E. coli (STEC) | Produce at least one Shiga toxin |
| Enterohaemorrhagic E. coli (EHEC) | Produce at least one Shiga toxin Induce AE lesions in the intestine Possess a virulence plasmid of about 60 MDa Cause haemorrhagic colitis and haemolytic uraemic syndrome |
| Enteroaggregative E. coli (EAEC) | Exhibit a typical aggregation when exposed to HEp-2 cells Some strains have the genes for the EAST1 enterotoxin Produce a cytotoxin |
| Enteroinvasive E. coli (EIEC) | Invade epithelial cells in a manner similar to Shigella Produce an enterotoxin |
| Diffusely adherent E. coli (DAEC) | Adhere to HEp-2 cells in a characteristic diffuse pattern Implicated in diarrhoea in young children |
The serogroups that are most frequently recovered from septicaemic calves and pigs have been identified.46, 53, 108
Necrotoxigenic E. coli (NTEC)
Necrotoxigenic E. coli produce one of two cytotoxic proteins CNF1 and CNF2.40, 144 These are large proteins which induce multinucleation in certain epithelial cell lines and necrosis in rabbit skin. Strains of E. coli which produce CNF1 (NTEC-1 strains) typically produce alpha haemolysin and P pili, whereas strains which produce CNF2 (NTEC-2 strains) typically produce F17-related pili. NTEC-1 strains have been reported in association with diarrhoea in calves, septicaemia in beef cattle, and lethal haemorrhagic colitis in adult cattle. NTEC-2 strains, especially of serogroup O78, are bovine septicaemic E. coli but only a small percentage of septicaemic O78 E. coli produce CNF2.111
Epidemiology
Whether or not disease is caused after an animal is infected with E. coli depends on a complex interaction between the susceptibility of the host, pathogenicity of the specific strain of the organism (see Aetiology and Pathogenesis), and environmental predisposing factors.
In all E. coli infections, except mastitis, disease develops only when the infecting E. coli possesses specific virulence factors which allow it to overcome host defence mechanisms. In enteric infections, the E. coli survive exposure to gastric acidity, adhere to the intestinal epithelium and multiply, and produce toxins or signals which alter the host physiology. If large numbers of E. coli are ingested then a sufficient number may pass through the stomach into the intestine even if a high percentage is killed in the stomach. Alternatively, the bacteria may cause disease in neonatal animals, before secretion of HCl in the stomach is well developed, or the bacteria may possess mechanisms for resisting exposure to low pH.
Extra-intestinal infections may occur through the oropharynx, the intestine, the urethra, the mammary gland, or the umbilicus.48, 53 The ability to evade the killing action of complement and to obtain iron from a low iron environment are two features often associated with E. coli that survive and multiply in the blood and tissues. In mastitis the bacteria act as opportunists and do not appear to require any special virulence attributes.
Host factors often play a critical role in the disease process. In order for pili to anchor bacteria to the intestine, the enterocytes must have receptors to which the pili can bind. Thus animals that fail to produce receptor molecules are resistant to colonization and disease by E. coli that carry the corresponding pilus.5, 49, 178 New-born and young animals are most susceptible to enteric colibacillosis. These animals are at a stage when the intestinal tract is designed to promote colonization and establishment of a normal flora, and pathogenic E. coli in the environment can take advantage of this state.168
Milk may contain specific protective antibodies as well as other molecules that inhibit colonization by pathogenic E. coli, and therefore weaning may render some animal species susceptible to E. coli diarrhoea.38, 152, 174, 186
Colostral antibodies are very important in the protection of new-born animals against both enteric and septicaemic infections.14, 15, 109, 152, 174, 186 Specific antibodies in the lumen of the intestine may protect against intestinal colonization, and antibodies in the blood will protect against septicaemic disease.48, 54
Colostrum should be ingested as soon after birth as possible, preferably within a few hours, in order to provide effective protection, as the enterocytes of neonates lose their ability to absorb antibodies in the colostrum as the animal gets older. The longer the interval between birth and the first ingestion of colostrum, the lower the amount of antibody that is absorbed which leads to inadequate protection.52, 109
The antibody classes in colostrum that are of particular importance in protection against E. coli infections are IgM and IgG in the case of colisepticaemia,14, 15, 53, 110 and IgA in enteric colibacillosis.6, 48
Infection with other infectious agents, such as rotavirus, coronavirus, Eimeria spp. or Cryptosporidium spp., may predispose young animals to enteric colibacillosis.2, 84, 171
As animals suffering from colibacillosis may excrete very large numbers of the pathogenic E. coli strain in almost pure culture, poor sanitary hygiene may result in massive faecal contamination of the external environment and the build-up of very large numbers of pathogenic E. coli. In addition, dampness of the environment plays an important role in the survival of E. coli and consequent spread of the infection.2
Pathogenic strains of E. coli to which animals on a farm have not previously been exposed may be introduced into a herd or flock by newly arrived animals or, conversely, newly arrived animals may become infected with strains with which they have not previously been in contact.66 These factors play a particularly significant role in calf-rearing units where the calves are purchased from many different sources. Stresses such as transportation and overcrowding also play major predisposing roles.2, 48
Pigs
Colibacillosis in pigs occurs in two distinct age groups: neonates and those in the immediate post-weaning period. The percentage of pathogenic strains of E. coli relative to nonpathogenic strains that is carried by the sow increases markedly at farrowing. This facilitates their transmission to piglets early in life. Whether or not neonatal colibacillosis develops in piglets which are exposed to pathogenic strains of E. coli depends to a large extent on the presence or absence of colostral- and lacteal-derived antibodies; other factors play a lesser role. On the other hand, stress plays an important predisposing role in the development of the disease in weaned piglets.
Piglets are exposed to many stress factors at weaning, including loss of maternal contact, movement to new pens, new pen-mates, overcrowding, erratic environmental temperature caused in part by lack of maternal warmth, and declining levels of lacteal-derived IgA. Weaning stress has been reported to cause a temporary increase in gastric pH, which results in decreased gastric bactericidal activity,159 but subsequent work did not support this finding.64 Changes in dietary composition at weaning or after transport to a different farm were believed to predispose piglets to colibacillosis.10, 11, 157, 167 It was reported that when piglets are fed a creep-feed while still suckling their dams, hypersensitivity to dietary antigens may develop which could predispose to enteric colibacillosis after weaning if they are fed a weaning diet containing the ingredients to which they developed sensitivity in the creep-feed.121, 122 Several studies have, however, failed to find supporting evidence.65, 67, 68, 101, 102
Highly nutritious diets that are rich in protein appear to favour colonization of pathogenic E. coli in the intestine of the weaned pig. The intestinal villi undergo changes at weaning, resulting in a period of malabsorption, which favours rapid growth of bacteria in the intestine.63, 68, 132 Supplementary feeding during the nursing period reduces the degree of malabsorption observed in the post-weaning period.132
The addition of antibiotics to feed enables antibioticresistant organisms in the intestine to multiply at the expense of the non-resistant ones. In the case of E. coli, pathogenic strains are often antibiotic-resistant.74
Certain serotypes of ETEC are associated with neonatal diarrhoea worldwide, but there are variations over time and from one country to another.135, 172, 184, 187 These E. coli are often non-haemolytic, mucoid, and K99-positive; or haemolytic, nonmucoid, and K88-positive. Escherichia coli that produce the 987P pilus may also cause neonatal diarrhoea in pigs. In weanedpigs, the E. coli that cause diarrhoea oroedemadisease are typically haemolytic. Strains implicated in post-weaning diarrhoea usually produce the K88ac (F4ac) or F18ac pili, whereas those associated with oedema disease usually produce the F18ab pili.136 The ETEC most frequently associated with diarrhoea are usually O8, O9, O20, O64, O138, O141, O147, O149 and O157.71, 72, 135, 172, 189
On analysis of the results of bacterial isolation from specimens of cases of colibacillosis in pigs submitted to the Onderstepoort Veterinary Institute in South Africa over a 13 year period (1987 to 1999), it was found that serotypes O149:K91 and O141:K85 comprised 58 per cent of isolates.75, 76
Strain O149:K91 was the predominant pathogenic serotype in pigs, accounting for 45 per cent of all pathogenic strains of E. coli that were isolated. Infection with this serotype is particulary common in the immediate post-weaning period (Table 150.2).74 Nearly 90 per cent of these isolates possessed the pilus attachment factor K88. Escherichia coli O141:K85 was the next most common isolate, of which about 40 per cent possessed the K88 antigen. This isolate accounted for about 13 per cent of the cases of colibacillosis in pigs. It is found in a wider age range of piglets, and was also isolated from aborted pig foetuses. All other pathogenic serotypes individually accounted for five per cent or less of the total number of isolates. The most common of these were E. coli O20, O8:K48, O2:K4, O9:K28 and O8:K87.
The two main syndromes seen in pigs are enteric colibacillosis (about 70 per cent of cases) and colisepticaemia (about 27,5 per cent). There has been a marked decrease in the prevalence of both syndromes in suckling piglets worldwide, including South Africa since 1981. This may be a reflection of the greater number of sows that are vaccinated (with consequent transfer of colostral antibodies to piglets), increased hybrid vigour due to the more frequent use of first-cross sows because they possess a greater non-specific disease resistance, and the improvement in farrowing housing.75
The oedema disease strains produce Stx2e and may also produce enterotoxins, most commonly one or both heatstable enterotoxins.1, 51, 184 The prevalence of oedema disease in South Africa is relatively low. Escherichia coli serotypes O139:K82, O138:K81 and O141:K85 are the most common causes of oedema disease worldwide.10 In South Africa serotypes O139:K82 and O138:K81 were usually associated with it before 1987. Serotypes which have been associated with oedema disease since 1987 are O149, O147 and O8, which do occur in South Africa, and O45, which as yet has not been isolated from pigs in this country.75, 76
Calves
Colibacillosis in calves is much more common in those fed milk surrogates than in ranched calves. Insufficient ingestion of colostrum soon after birth appears to be the most important predisposing factor. It occurs mostly in spring.
In South Africa the most common pathogenic E. coli serotypes isolated from specimens from new-born calves submitted to the Onderstepoort Veterinary Institute, are O78:K80, O9:K30, O119:K69, O20 and O9:K28, comprising 70 per cent of all serotypes isolated (Table 150.3).75
Table 150.2 The number of isolates of Escherichia coli of various serotypes associated with disease in piglets of different age groups over the 13-year period 1987 to 1999 in South Africa75
| AGE OF PIGS | O149:K91:K88 | O141:K85:K88 | OTHER | TOTAL (%) |
|---|---|---|---|---|
| Foetus | 1 | 5 | 5 | 11 (1,6) |
| Pre-weaning | 93 | 22 | 127 | 242 (34,4) |
| Post-weaning | 193 | 57 | 121 | 371 (52,8) |
| Adults | 31 | 8 | 40 | 79 (11,2) |
| Total | 318 (45,2%) | 92 (13,1%) | 293 (41,7%) | 703 |
Table 150.3 Common Escherichia coli serotypes isolated from calves over the 13-year period 1987 to 1999 in South Africa75
| SEROTYPES OF E. COLI | NUMBER ISOLATED | ||||
|---|---|---|---|---|---|
| Septicaemia | Abortion | Other | Total | ||
| O78:K80 | 75 | 30 | 4 | 3 | 111 |
| O119:K69 | 50 | 7 | 0 | 1 | 58 |
| O9:K30 | 40 | 13 | 1 | 2 | 55 |
| O26:K60 | 37 | 6 | 0 | 2 | 45 |
| O20 | 24 | 15 | 1 | 1 | 42 |
| O9:K28 | 18 | 10 | 2 | 1 | 29 |
Table 150.4 Common Escherichia coli serotypes isolated from sheep and goats over the 13-year period 1987 to 1999 in South Africa75
| SEROTYPE | SHEEP | GOATS | TOTAL |
|---|---|---|---|
| O78:K80 | 33 | 16 | 49 |
| O113:K75 | 12 | 4 | 16 |
| O8:K50 | 14 | 1 | 15 |
| O137:K79 | 6 | 7 | 13 |
| O20 | 6 | 5 | 11 |
Escherichia coli serotypes such as O78:K80, O26:K60 and O111:K58 occur in a much wider age range calves than O9:K30. Some are pathogenic not only for neonatal animals, but also for calves up to one month old, and may also cause abortions in cattle.75 In calves more than two weeks old, E. coli infections often occur in association with other diseases, such as paratyphoid, chlamydiosis and pasteurellosis. Poor housing conditions and nutrition enhance their pathogenicity.
Lambs and kids
Colisepticaemia occurs much more commonly than enteric colibacillosis in intensively raised lambs and kids.75 More cases in lambs occur in autumn than in spring. Escherichia coli O78:K80 and O113:K75 serotypes were most commonly isolated from specimens of affected lambs and kids in South Africa over a 13-year period (Table 150.4). The adhesive factor K99 rarely occurs in serotypes of E. coli of small stock in South Africa, and was only present in a limited number of strains of the O9:K30 and O141:K85 serotypes. This is probably the main reason for the rare occurrence of enteric colibacillosis in small stock in this country.75
Foals
Escherichia coli is a rare cause of endometritis and abortion in mares and of colisepticaemia in foals.75, 78, 85, 151 Worldwide, ETEC infections are implicated only occasionally as a cause of disease in foals.82, 83, 85, 151
Escherichia coli serotype O78:K80, and serogroups O113 and O9 group have been isolated from diseased foals in South Africa but, in general, E. coli infection tends to complicate diseases caused by infection with Salmonella, Pasteurella or Actinobacillus spp.75 The ETEC serotypes isolated from horses carry F41, K88 (F4), K99 (F5) and 987P (F6), inconsistently.82
Pathogenesis
Enterotoxigenic E. coli (ETEC) infections
Escherichia coli is part of the normal intestinal flora, and pathogenic strains form only a small part of the total E. coli population of adult animals. It is usually transmitted by the faecal-oral route, and is one of the first organisms to establish itself in the large intestine soon after birth, with the small intestine being colonized subsequently. Once established, and irrespective of age, few E. coli are present in the cranial part of the small intestine while relatively large numbers are present in its caudal part and in the large intestine.91, 149, 168
Several factors affect the success of establishment of ETEC in the intestinal tract.4, 9, 33, 38, 63, 65, 149 Ingestion of large numbers of ETEC increases their chances of passage through the stomach and survival in the intestine. Receptors for specific pili on the small intestinal epithelial cells and overlying mucus determine susceptibility or resistance of pigs to ETEC with specific pili, the area of intestine that is colonized, and the age-related susceptibility to certain ETEC. Receptors for K88 pili are most concentrated in the cranial small intestine of pigs, hence this is the area of the intestine that is most heavily colonized with K88-positive ETEC. K99-positive ETEC affect young pigs, calves, and lambs but not older ones because there is an age-related decrease in the concentration of small intestinal receptors for K99 pili. In contrast, K88-positive E. coli affects both neonatal and weaned pigs, because receptors for these pili persist in older animals.
The presence or absence of specific antibodies in the intestine is a major factor in determining whether disease will develop following exposure to ETEC.3, 162, 167, 183 Other compounds in milk or the feed may also impact on growth and adherence of bacteria as well as absorption from the intestine.4, 10, 11, 38, 141, 167
The pathogenesis of ETEC infections, in all species and age groups is similar. Ingested bacteria pass through the stomach in sufficient numbers to permit them to establish in the small intestine.
In order to protect themselves against rapid elimination from the small intestine, ETEC adhere to specific receptors in the small intestine. The adherence is mediated by protein adhesins carried on large numbers of pili which project from the bacterial surface. The polysaccharide capsule appears to promote colonization in calf and porcine ETEC, possibly by strengthening the attachment of piliated bacteria to the intestine. Adherent bacteria multiply and reach large numbers in the intestine. For example, the number of E. coli in the mid-jejunum of a healthy calf is in the order of 104 per gram of intestinal content, but this number rises to approximately 109 per gram in a calf with enteric colibacillosis.91 Enterotoxigenic E. coli that have colonized the small intestine produce one or more enterotoxins, which cause accumulation of fluid and electrolytes in the intestine. Diarrhoea develops because the absorptive capacity of the intestine is exceded. Loss of electrolytes and water from the intestine leads to dehydration, metabolic acidosis, and possibly death.
Several adhesins have been identified as playing roles in virulence of ETEC for livestock.37, 87, 125, 127, 133, 135, 189 These adhesins are carried by pili which are short proteinaceous filaments projecting from the surface of bacteria and are not involved in movement or the transfer of nucleic acid. Typically, pili are more numerous than flagella (100 to 500 per cell) and are much thinner (2 to 8 nm in diameter). The major pili that are involved in colonization of the intestine by ETEC are called K88 (F4), K99 (F5), 987P (F6), F41, F107 and F18. Various combinations of these pili are sometimes associated with a single strain of ETEC. Typically, the genes for synthesis of the pili are temperature regulated and are activated at 37 °C but not at 18 °C.
Both K88 and K99 pili carry K antigen designations because they were not initially recognized as fibrillar structures. They have been re-named F4 and F5, respectively, but the names K88 and K99 are deeply entrenched in the literature. There are three major antigenic types of K88—K88ab, K88ac, and K88ad. K88 pili are produced principally in association with O8, O45, O138, O141, O147, O149, and O157.37, 71, 72, 125, 135 Strains of O8 may possess K88ab, K88ac or K88ad pili; strains of O141 may produce K88ab or K88ac; and strains of other O groups usually express K88ac. Typically, K88-positive E. coli produce heat-labile enterotoxin (LT) alone or in combination with heat-stable enterotoxins (ST)a and/or b). The K88 pili are encoded by genes on a plasmid.
Five porcine phenotypes have been described on the basis of susceptibility of their intestinal epithelial cells to attachment by K88ab, K88ac and K88ad pili.5, 18, 49 K88 pili are important in attachment of bacteria to the intestine of both neonatal and weaned pigs.135, 161, 169, 170 The mucus covering the intestinal mucosa of older pigs has a much higher concentration of K88 receptors and is believed to protect the underlying brush border, but there appears to be a temporary breakdown in protection after weaning.31, 37
K99 (F5) pili occur on bovine as well as porcine ETEC. They show a decrease in binding as the intestinal epithelium ages, apparently due to an age-dependent decrease or masking of receptors. K99 pili are associated with strains of O groups 8, 9, 20, 64, and 101, which produce a K antigen with abundant capsular polysaccharide. Bovine K99-positive ETEC produce STa as the only enterotoxin, but porcine ones produce STa, STb, or both STs.72, 130 The amino acid alanine (or leucine) induces a repression of the biosynthesis of the K99 pili, and growth of the bacteria on special media is often required for good expression of the pili.37 Genes for K99 are carried on plasmids of various sizes. The best characterized plasmids are large, nonconjugative 50 MDa plasmids, but plasmids which encode ST, colicin, and drug resistance also encode the K99 pili.74
Enterotoxigenic E. coli strains of serogroups O9 and O101 produce the adhesive antigen designated F41.37, 131 Typically, F41 and K99 antigens are found together on the same strains of E. coli. However, F41+, K99− strains of E. coli of O serogroup 101 have been reported in association with diarrhoeal disease in pigs.131, 175, 189
The 987P (F6) pili are found on certain porcine ETEC.37 They are difficult to detect on bacteria grown in vitro and it may be necessary to passage a culture in vivo. These pili are approximately 7 nm in diameter and have a rigid appearance. The genes for 987P are both plasmid-borne and chromosomal and both copies of the genes need to interact to result in assembly of pili. Escherichia coli with this pilus antigen belong to a large number of O groups and produce one or both STs.37 Age-related resistance to colonization of the pig’s intestine occurs for 987P-positive E. coli by a mechanism similar to that described for K88-positive ETEC.33
The F18 pili have a shared ‘a’ antigen and distinct ‘b’ and ‘c’ antigens that give rise to F18ab and F18ac types.136, 154, 185 They require special growth conditions to facilitate their expression in vitro.185 The O139 oedema disease strains of E. coli usually produce F18ab pili whereas post-weaning diarrhoea strains of O138 and O141 produce F18ac pili and may or may not have the ability to cause oedema disease.87, 136, 185 In non-O139 E. coli the genes for the F18 pili are often on a plasmid which also carries genes for alpha haemolysin. F18 pili adhere to the intestine of older but not younger pigs.134
Enterotoxigenic E. coli produce two major types of toxins, heat labile (LT) and heat stable (ST).34, 60, 165, 166 A single type of LT, a toxin that is very similar to cholera toxin, and two distinctly different heat-stable enterotoxins, STa (STI) and STb (STII) are produced by ETEC implicated in disease. Each toxin may be produced alone or in combination with the others. The genes for enterotoxins are carried on plasmids. In southern Africa, LT production is mainly associated with E. coli O149:K91.76 Both types of enterotoxin induce changes in the intestinal epithelial cells that result in hypersecretion of fluids and electrolytes into the lumen of the intestine. A third heat-stable enterotoxin, called enteroaggregative E. coli stable toxin 1 is a 38-amino-acid polypeptide which is similar in its mode of action to STa.
Genes for this toxin have been found on a plasmid in porcine K88- positive ETEC,190 and on 66 per cent of CNF1 positive E. coli from cattle with septicaemia and diarrhoea.7
Heat-labile enterotoxin is composed of one A subunit of 28 kDa and five B subunits of 11,5 kDa.55, 56, 60, 137 The A subunit possesses enzymatic activity which is responsible for the toxicity, while the B subunits mediate binding of the toxin to receptors. Proteolytic enzymes in the intestine may nick the A subunit to produce a small A2 fragment and a larger A1 fragment joined by a disulphide bond. The B subunits bind primarily to ganglioside GM1 receptor molecules on enterocytes of the small intestine and mediate internalization of the toxin molecule in an endocytic vacuole, which is routed through the Golgi apparatus. Subsequently, the A1 fragment is released by reduction of the disulphide bond and escapes from the vacuole. The A1 fragment catalyzes the transfer of adenosine diphosphate-ribose from nicotinamide adenine dinucleotide to a regulator of adenylate cyclase, causing persistent activation of adenylate cyclase. This results in a marked increase in the concentration of cyclic adenosine monophosphate (cAMP) in the cells, and leads to activation of protein kinase A and excessive phosphorylation of the cystic fibrosis transmembrane channel. These changes cause hypersecretion of Cl− ions in the crypt cells and inhibition of absorption of Na+ and Cl− ions in the cells of the villus tip. Both activities cause excessive amounts of electrolytes and water to accumulate in the lumen of the intestine, resulting in diarrhoea. Alternative pathways involving prostaglandins, the enteric nervous system, and cytokines may also contribute to hypersecretion of electrolytes.
Heat-stable enterotoxin, STa, is an 18- or 19-amino-acid peptide that is produced by some ETEC.60 It binds to the receptor portion of particulate guanylate cyclase C on the surface of intestinal epithelial cells. Binding activates guanylate cyclase C which converts guanylate triphosphate to cyclic guanylate monophosphate (cGMP). The elevated levels of cGMP cause an efflux of Na+ ions and a reduction in absorption of Cl− ions, both actions contributing to excessive fluid in the intestine and diarrhoea. In South Africa, production of STa is associated with ETEC of serogroups O20, O154, O9:K30 and O9:K35.75
Heat-stable enterotoxin, STb, is a 48-amino-acid peptide that is produced by certain porcine ETEC.42 It appears to bind to sulfatide in the intestinal mucosa155 but little is known about its mode of action. Following absorption of the toxin there is release of 5-hydroxytryptamine and production of prostaglandin E2, suggesting that the toxin acts through the enteric nervous system.
Shiga toxin-producing E. coli (STEC) infections
Infection by STEC, like those by ETEC, involve intestinal colonization followed by the alterations in host fluid metabolism described for ETEC. The pathogenesis of STEC disease in calves, however, involves colonization by quite different processes and mechanisms that result in intestinal fluid accumulation in the intestine that do not involve enterotoxins.
No pilus adhesins have been associated with colonization by bovine STEC, which adhere very closely to altered host epithelial cells. Shiga toxin that is produced by STEC appears to have its major effect after it is absorbed from the intestine, and the disease syndrome that develops is markedly influenced by the extent to which the toxin is absorbed from the intestine and by the organs which have receptors for the toxin.137
Fluid loss appears to be due to processes activated by signal transduction in enterocytes and to loss of the absorptive surface due to effacement of the microvilli. Haemorrhage and escape of fluids are attributed to the action of Stx which is absorbed locally and causes vascular damage in the terminal ileum, caecum and colon. Signal transduction events triggered by the bacterial molecules that are ‘injected’ into the host epithelial cells cause hypersecretion of Cl− ions and also contribute to diarrhoea. In calves, there is no indication that Stx is absorbed from the intestine in quantities sufficient to have a recognizable systemic effect.
The pathogenesis of oedema disease in pigs involves colonization similar to that occurring in ETEC infections, but diarrhoea is not a feature of the disease, which is largely the result of a toxaemia due to absorbed Stx2e.1, 10, 11, 88, 100, 104, 140 The STEC that cause oedema disease produce alpha haemolysin, whereas those that cause disease in calves usually produce a related toxin called enterohaemolysin or EHEC haemolysin. A role in disease has not been demonstrated for these haemolysins.
Shiga toxin-producing E. coli are carried as part of the normal rumen and intestinal flora of cattle and as part of the intestinal flora of pigs.51, 116 However, whereas in healthy carrier pigs STEC constitute a minor part of the faecal E. coli flora, in animals with oedema disease they are the dominant or the only E. coli recovered from faecal culture.172
In calves STEC in the environment are ingested and pass through the stomach into the intestine, where they adhere to epithelial cells and colonize.92, 93, 188 Colonization occurs primarily in the colon and caecum, where attaching-andeffacing (AE) lesions develop.35, 93, 188 A chromosomal locus for enterocyte effacement (LEE) is necessary for development of the AE lesions.114, 115 LEE-encoded proteins include proteins of a type III secretion system, an adhesin (intimin) and its receptor, and signal transducing proteins. Two LEEencoded molecules known to be important in development of the AE lesions are called EspB (E. coli secreted protein B) and Tir (translocated intimin receptor).137 Shiga toxinproducing E. coli in close proximity to intestinal epithelial cells transfer the intimin receptor and signal transducing proteins to the host cell. EspB initiates a signalling cascade in the host epithelial cell which results in cytoskeletal rearrangements and the secretion of fluid from the cell.
The rearrangements include loss (effacement) of the villi on the cell surface and formation of pedestals with accumulated actin at the base. Tir becomes associated with the surface of the host epithelial cell membrane and acts as the receptor for intimin or Eae (E. coli attaching and effacing) protein. Variations in intimin are associated with the host species colonized by STEC.147 The end result is that the bacteria adhere very closely to the surface of epithelial cells which have lost their microvilli and have undergone marked morphologic alterations associated with rearrangement of the actin cytoskleton. Intimin binds to the receptor (Tir) to form a very intimate adherence.
The mechanism of attachment of STEC to the intestine of pigs is similar to that described for ETEC and does not involve damage or alterations to the intestine.12, 120 Whereas bovine STEC have evolved from EPEC, with which they share a colonization mechanism, oedema disease strains of E. coli have evolved from ETEC and have the same mechanism of colonization. The pili which mediate colonization by oedema disease strains of E. coli are F18ab.136, 154, 184
Shiga toxin-producing E. coli are characterized by their ability to produce at least one Stx.99, 137 There are two categories of Stx, known as Stx1 and Stx2. Stx1 is almost identical to Shiga toxin produced by Shigella dysenteriae type 1 and is neutralized by antiserum produced against this toxin. Stx2 toxins are not neutralized by anti-Shiga toxin serum. Whereas Stx1 is highly conserved, there are several variants of Stx2, including Stx2e which is responsible for oedema disease of pigs. Ruminants, especially cattle, are a major source STEC which cause disease in humans.116, 137 The genes for Stx1 and Stx2 are carried on temperate bacteriophages which may play a role in regulation of toxin synthesis as well as in release of toxin through phage-mediated cell lysis.
Shiga toxins have one A subunit (32 kDa) associated with a pentamer of B subunits (7,7 kDa). The A subunit is converted to A1 (N-terminal 27 kDa) and A2 (C-terminal 4 kDa) fragments by proteolysis and reduction.137, 142 The B subunits bind to a glycolipid receptor (either globotriaosyl ceramide 3, or globotetraosyl ceramide 4) in the membrane of epithelial or endothelial cells.24, 137 Following binding, the toxin is internalized by receptor mediated endocytosis and is transported to the Golgi apparatus and then to the endoplasmic reticulum, where release of the A subunit allows interaction with ribosomal RNA. The N-glycanase activity of the A1 fragment cleaves a specific adenine residue from 28S ribosomal RNA, thereby inhibiting protein synthesis. In intestinal epithelial cells this action results in increased fluid secretion. In blood vessels, damage to vascular endothelial cells results in leakage of fluids and red blood cells into the tissues, and in intravascular coagulation and thrombosis.
Shiga toxin 2e (Stx2e) produced by oedema disease E. coli (also known as oedema disease toxin or oedema disease principle), 30, 41, 51, 58, 119 binds preferentially to globotetraosyl ceramide. The purified toxin induces the signs and lesions of oedema disease in pigs injected intravenously with as little as three nanograms of toxin per kg bodyweight.113 In pigs that die of the natural disease, the toxin can be demonstrated in the intestinal contents172 and damage to vascular endothelium is observed in target tissues.10, 100, 104
However, the toxin does not appear to be absorbed from the intestine under normal conditions179 and it is likely that unusual conditions in the intestine are required to permit absorption in those pigs that develop disease. Antibodies against the inactivated toxin neutralizes the toxicity of Stx2.112
Alpha haemolysin is a protein cytolysin which is a virulence factor for E. coli that cause urinary tract infection in humans. This cytolysin is a 110 kDa member of the RTX (repeats in toxin) family of pore-forming cytotoxins, which damage membranes of a variety of cells. Porcine ETEC and oedema disease strains of E. coli often produce alpha haemolysin, but a role in diarrhoea or oedema disease has not been demonstrated.10, 64, 170 Antibodies to the haemolysin are produced in pigs that develop oedema disease. Elimination of the plasmid with the genes which encode the haemolysin, however, did not prevent the bacteria from causing the disease under experimental conditions.170
Enterohaemorrhagic E. coli haemolysin (enterohaemolysin) is another RTX toxin which is produced by some STEC.137 The genes are found on a large virulence-associated plasmid and the EHEC haemolysin operon has an organization which is similar to that for alpha haemolysin. The EHEC haemolysin has weak activity and its haemolytic activity is not detected unless washed red blood cells are used in agar media.17
Disease in calves caused by STEC is usually sporadic and there is little information on factors that affect the disease process.116 Most adult cattle have had exposure to STEC and a high proportion (78,4 per cent) contain neutralizing antibodies in their serum.22 These are probably transferred via the colostrum to the intestine of calves. Furthermore, disease only infrequently occurs in calves younger than two weeks of age. These observations suggest that maternally derived antibodies ameliorate the course of disease. The disease has also been reported in goats.43
The pig’s diet influences colonization of the intestine by STEC. Highly nutritious diets predispose to disease whereas poorly nutritious diets that are high in fibre exert a protective effect.9, 10, 88, 167, 172
Enteropathogenic E. coli (EPEC) infections
These infections involve colonization and formation of the AE lesions in a manner similar to those described for STEC. In cattle, potential EPEC have been identified almost exclusively in the context of attaching and effacing E. coli (AEEC) or eae-positive E. coli which are negative in tests for Shiga toxins or their genes.28, 116, 143, 156 However, E. coli that are positive for eae and negative for stx may be EPEC, STEC which have lost their stx genes, or neither of the two.
Tests for the presence of the EHEC virulence plasmid and the EAF plasmid may help in categorizing such isolates.156
Enteropathogenic E. coli are implicated in diarrhoea in weaned pigs, and the serogroup that has been best characterized as a porcine EPEC is O45:K‘E65’.191 These E. coli possess the LEE and induce AE lesions primarily in the small intestine. However, porcine EPEC lack a counterpart to the enteropathogenic E. coli adherence factor plasmid and bundle-forming pili, suggesting that there are differences from human EPEC in generation of the AE lesions.191
Colisepticaemia
In most countries septicaemia is encountered only infrequently in pigs, but ETEC are often recovered from organs of pigs that have died from diarrhoea and dehydration.46, 172 This phenomenon may represent failure of moribund pigs to clear from their circulation ETEC that translocate from the intestine. In the septicaemic disease, bacteria with poorly defined virulence factors invade through the intestine or the umbilicus and multiply in the blood stream, most commonly in new-born pigs. Bacterial properties identified in some of these invasive E. coli include resistance to the killing action of complement in serum and ability to obtain iron from low iron environments. Typically, septicaemic E. coli lack the pili and enterotoxins which characterize the enteric pathogens. Serogroup O78:K80 has been associated with septicaemia in pigs in several countries, including South Africa. Escherichia coli bearing F165 pili and belonging to several O serogroups have been implicated in septicaemia and diarrhoea in pigs.138, 139
Septicaemia occurs in calves which have deficient levels of serum gammaglobulin and have been infected with E. coli capable of causing septicaemia.14, 52, 96 Affected calves may be agammaglobulinaemic or hypogammaglobulinaemic, a state usually caused by failure to ingest colostrum early enough or in sufficient quantity.14 The protective effect of gammaglobulin is not dependent on specific antibodies against the septicaemic E. coli.
Virulence attributes of bovine septicaemic E. coli have not been well identified. The major factors associated with these E. coli are resistance to serum bactericidal activity and possession of an aerobactin iron capture system which allows the bacteria to sequester iron in the low iron environments found in serum and tissues.53 TheOKserogroup most frequently implicated in bovine septicaemia is O78:K80.48
Escherichia coli multiply in the blood and cause the affected calves to develop shock, attributable to endotoxaemia. Such calves may collapse and die acutely, or the illness may run a less acute course, with sequelae such as polyarthritis and meningoencephalitis.59, 160 Many of these calves develop diarrhoea, in some cases attributable to endotoxaemia and in others due to intercurrent infection with ETEC.
Lipopolysaccharide (LPS), also referred to as endotoxin, is present in the cell wall of Gram-negative bacteria. The LPS molecule is composed of lipid A, a central core, and repeating repeating carbohydrate units which constitute the O antigen.
Toxicity is associated with lipid A, but the composition and length of the O-antigenic chain influence interactions of the bacterium with host defence systems. Minute quantities of LPS that are absorbed from the intestine into the circulation are detoxified by the liver and cells of the reticuloendothelial system. Larger amounts of LPS may be found in the circulation in cases with septicaemia and in those where damage to the intestine permits excessive uptake of LPS from the intestine. Fever, disseminated intravascular coagulation and shock can be caused by LPS injected into animals and in cases suffering from Gram-negative bacterial septicaemia.70
Lipopolysaccharide is a signal for the host to mobilize a range of responses that are essential for defence against Gram-negative bacteria. Its activation of the alternate complement pathway can result in lysis of Gram-negative bacteria, enhancement of phagocytosis through opsonization, increase in vascular permeability, attraction of leukocytes, and release of histamines and of lysosomal enzymes of granulocytes. The effects of LPS are largely due to activation of mediator cells, principally macrophages. Activation of these cells leads to production of various effector molecules including leukotrienes, prostaglandins, and cytokines. A moderate response to LPS is helpful to the host but a strong response can lead to vasodilation, fall in blood pressure, disseminated intravascular coagulation, and shock.
The O-antigen capsule called K‘V165’ is responsible for resistance to the bactericidal effects of complement as well as for resistance to phagocytosis by porcine polymorphonuclear leukocytes.45, 138, 139 Other capsular polysaccharides may endow E. coli with serum resistance by presenting a surface which does not promote binding of complement component C3b. Certain polysaccharides of LPS may have a similar effect. Many strains of septicaemic E. coli possess an F plasmid which encodes colicin V and a number of proteins associated with virulence.19, 164 One of these is the TraT outer membrane protein, which may act by interfering with the insertion of the C5-C9 membrane attack complex into the bacterial outer membrane.
The presence of the aerobactin-iron-uptake system is a common feature of septicaemic E. coli. This is not surprising since continued bacterial growth in blood and tissues requires that the bacteria acquire iron from host glycoproteins to which iron is tightly bound. This iron uptake system allows the E. coli to secrete a high affinity hydroxamate iron chelator known as aerobactin, which is particularly well suited to operate in systemic infections.19 The genes for this system are also present on the colicin V plasmid in septicaemic E. coli.
Cytotoxic necrotizing factor (CNF) 1 and CNF2 are E. coli protein toxins of 110 kDa and 115 kDa, respectively.40 The toxins are highly lethal for mice, cause necrosis in rabbit skin, and induce multinucleation and polymerization of actin in certain cultured cells. CNF1 has a more powerful ability to induce multinucleation and less ability to cause dermonecrosis than CNF2.
The genes for CNF1 are carried on the chromosome while those for CNF2 are carried on a transmissible plasmid called pVir, but the genes have 85 per cent amino acid sequence identity. Both toxins modify the guanine triphosphatase (GTPase)-binding protein Rho by deamidation which results in a decrease in intrinsic GTPase activity, leading to constitutive activation of Rho, and reorganization of actin stress fibres.173
Strains of E. coli which produce CNF1 usually produce alpha haemolysin and have been recovered from a variety of infections in humans and animals as well as from healthy animals. Genes for P pili, CNF1 and alpha haemolysin are sometimes clustered as a mobile genetic element. CNF2-producing E. coli have been isolated from healthy animals as well as from calves and pigs with septicaemia or diarrhoea.
Clinical signs
Enteric diseases due to E. coli
Enteric colibacillosis in pigs occurs predominantly in the neonatal period and one to two weeks after weaning.20, 135, 172, 183 In new-born piglets, the incubation period may be as short as two to three hours. Affected piglets may be found dead without clinical signs having been noticed, or they may be moribund with or without evidence of diarrhoea. If diarrhoea is present, the faeces are watery and whitish to yellowish. In the longer-standing disease, they may show weakness, anaemia due to reduced absorption of nutrients (especially iron), dehydration and loss of condition. The mortality rate is highly variable but may approach 100 per cent, particularly in very young piglets.
In post-weaning diarrhoea caused by E. coli-affected pigs commonly show diarrhoea, but they may be found dead without having shown signs of illness.64, 172, 177 The faeces are usually watery and greyish or brownish, and may be haemorrhagic.153 The diarrhoea often resolves within a few days, but it may persist and result in death from dehydration or in chronic enteritis and stunting. The morbidity rate may be as high as 50 per cent and the mortality rate is usually less than 10 per cent.
Oedema disease occurs sporadically, is stress-associated, and most commonly affects recently weaned pigs in good condition.10, 44, 88, 163, 172 One or more pigs in a group may be found dead without clinical signs having been noticed, while some of the others show nervous signs such as opisthotonus, incoordination, paddling, dullness, apparent blindness and head-pressing.
The subcutaneous tissues of the head, especially of the eyelids (Figure 150.1), nose and ears are oedematous; and affected animals often have a hoarse, squeaky voice attributable to laryngeal oedema. The neurological signs may be preceded by diarrhoea if the E. coli strain associated with the disease produces enterotoxin. Respiratory distress is a common terminal sign. Death usually occurs within four to 36 hours of the onset of clinical signs. The morbidity rate in a herd is usually less than 15 per cent, but more than 50 per cent of affected pigs will probably die.
Enteric colibacillosis caused by ETEC in calves occurs primarily in the first two to three days of life.2, 20 In the peracute disease, calves may die before enteritis is evident. Acutely affected animals are listless, dull, disinclined to drink, and usually show a profuse, watery diarrhoea which is whitish-yellow or greyish. Depending on the severity of diarrhoea, calves show variable degrees of dehydration. The morbidity and mortality rates may be high.
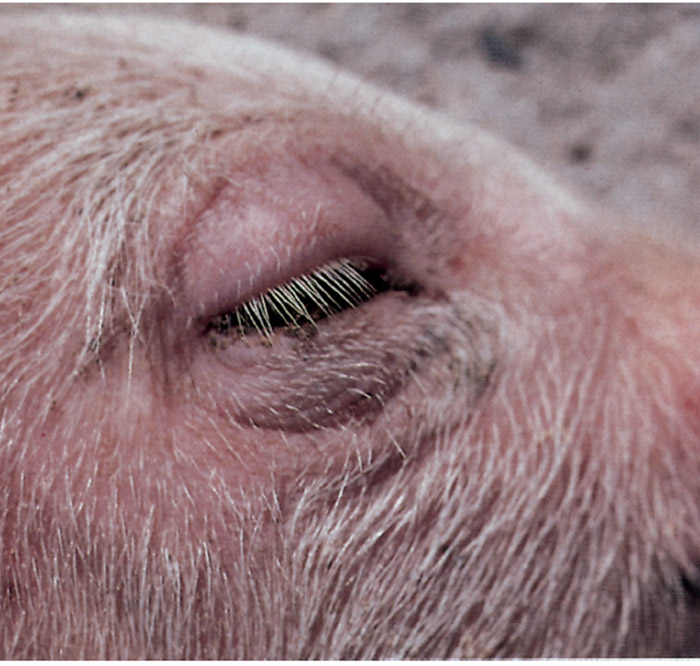
Strains of AEEC may cause disease in calves of up to four months of age, but calves less than three weeks old are usually affected.26–29, 92, 93 These strains may be EPEC or STEC, and the diarrhoea, often containing blood or mucus, that they induce is the predominant clinical sign. Affected calves may become dehydrated and refuse to drink. The disease may be mild or severe and life-threatening.
In calves more than two weeks of age E. coli has been isolated in association with other diseases such as paratyphoid, chlamydiosis or pasteurellosis and it is often difficult to assess the role that E. coli played (if any) in the pathogenesis of these diseases.75
Occasionally, calves will show clinical signs of dysentery associated with certain serotypes of STEC but the factors which predispose to such disease are not known.26, 62 The STEC which cause dysenteric disease in calves belong predominantly to serogroups O5:H−, O26:K60, O111:H11 and O111:H−, and have characteristics of human EHEC.29, 116, 148, 150, 181, 182 There is epidemiological evidence that STEC, especially those which produce Stx1, are implicated in a watery diarrhoea syndrome in calves, but the data vary considerably from one study to another.181, 182, 188 In parts of Europe, strains of O118 are closely associated with calf diarrhoea.182
Other diarrhoea-inducing agents such as ETEC, rotavirus, coronavirus, bovine viral diarrhoea virus or coccidia may be present in calves with STEC diarrhoea.93, 176 Calves affected with STEC tend to be older than those that suffer from ETEC infections.
Bovine EPEC have not been well characterized and have been identified largely as Stx-negative attaching-andeffacing E. coli.
Anti-bacterial antibodies in the intestine of the calf may ameliorate the course of disease or prevent disease. Intercurrent infection with other enteric pathogens, notably rotavirus, may increase the susceptibility of calves to ETEC infection.171
Septicaemic disease in pigs and calves
The course of disease is usually acute, ranging from one to three days.46 Pigs affected by colisepticaemia are febrile and listless but soon become recumbent and unconscious, or may show paddling limb movements before death. Calves are initially depressed and feverish, but the temperature drops rapidly as they become moribund. They may show incoordination, opisthotonus, recumbency and leg-paddling movements.20
Colibacillosis in lambs and kids
Both enteric and septicaemic colibacillosis (Table 150.4) occur primarily in intensively reared lambs and kids. The clinical signs are similar to those described in piglets and calves.
Colibacillosis in foals
The ETEC serotypes isolated from horses sometimes carry F41, K88 (F4), K99 (F5) or 987P (F6) pili.81–83, 85 The clinical signs in foals affected by enteric colibacillosis or colisepticaemia are similar to those in the other animal species. Stx-positive and eae-positive E. coli have been isolated from foals with diarrhoea.81
Other diseases
Apart from enteric colibacillosis, colisepticaemia and oedema disease, E. coli may also occasionally cause abortion, metritis, cystitis, mastitis and agalactia.20 Abortion due to E. coli in cattle is rare and usually occurs at a late stage of gestation. Escherichia coli may be isolated in pure culture from all organs of such aborted foetuses. In South Africa, E. coli O78:K80 is the strain which is most commonly associated with abortion in cattle.75
Escherichia coli is the cause of sporadic cases of acute mastitis in cows in early lactation. The serotypes involved are very diverse as they are generally of faecal origin and act as opportunistic pathogens. Escherichia coli from the environment enter the teat duct and make their way into the gland cistern, where they multiply and induce a marked inflammatory response. Absorption of endotoxin produced by the E. coli can lead to severe toxaemia and death.
Escherichia coli mastitis also occurs in sows.13 Endometritis and abortion in mares have also been reported, but are rare.
Pathology
Animals that are affected by enteric colibacillosis due to ETEC are dehydrated and their hindquarters are soiled with faeces as a result of diarrhoea. Necropsy usually reveals a fluid-filled and congested small intestine, and oedematous and congested mesenteric lymph nodes.97
In the acute septicaemic disease, gross lesions include: generalized congestion and the presence of petechiae and ecchymoses on serosal surfaces; congestion of the meninges, lungs, the mucosa of the stomach and/or intestine; and oedema of the meninges, and/or lungs.97 In less acute cases, there may be evidence of organ or tissue localization of the bacterium, with lesions such as fibrinopurulent pleuritis, pericarditis, peritonitis, polyarthritis, pneumonia and meningitis.59 Calves may also show a meningoventriculitis. In some animals, fibrinopurulent omphalophlebitis may be evident. In calves surviving the septicaemic disease, ‘white-spotted kidneys’ due to focal disseminated interstitial nephritis have been attributed to E. coli infection.
In calves that have died from STEC infection, AE lesions may be demonstrated in the intestine. Light microscopy shows multiple foci of attached bacteria often aggregated in micocolonies and commonly in association with an irregular appearance to the epithelial surface.26, 27, 62, 148, 150, 188 Enterocyte necrosis and stunting of villi are frequently observed. Adherent bacteria are much clearer in sections stained with Giemsa stain than in those stained with haematoxylin and eosin. The classical attachment of the bacteria to epithelial cells and effacement of microvilli are seen by transmission electron microscopy. The STEC are closely adherent to the epithelium, which is denuded of microvilli. There is a dense accumulation of polymerised actin in the epithelium beneath the attached bacterium and the architecture of the epithelial cell is often changed to form a pedestal on which the bacterium lies.
Pigs that have died from oedema disease are generally in good condition.10, 11, 97, 172 The severity and distribution of the lesions vary considerably from case to case; in some they are quite obvious, while in others they must be actively sought during a necropsy. Oedema and congestion, with or without haemorrhage, may be present in some or all of the following tissues: the subcutaneous tissues of the eye-lids (Figures 150.1), ears, forehead, jowl and ventral abdomen; the mesocolon; the submucosa of the greater curvature of the stomach, spiral colon, larynx and oesophagus; and the mesenteric and superficial lymph nodes. There may be an increased amount of peritoneal fluid which contains strands of fibrin.
Histopathologically, apart from the oedema, the most significant lesion of oedema disease is a degenerative angiopathy affecting the small arteries and arterioles in many organs and tissues.10, 100, 104 Initial perivascular oedema is followed by necrosis of smooth muscle cells of the tunica media, which then undergoes fibrinoid necrosis. This is followed by proliferation of cells in the tunica adventitia. Swelling of endothelial cells has also been observed, but thrombosis is not a prominent feature. These vascular changes may be difficult to discern in acute cases. In animals which survive for several days, foci of encephalomalacia are often present in the brain stem, frequently with a bilaterally symmetrical distribution, as well as cerebral vascular lesions.
Diagnosis
The clinical signs of enteric colibacillosis, namely depression, anorexia, diarrhoea and dehydration, are similar to those of infections caused by other enteric pathogens, such as rotavirus, coronavirus, Salmonella, and Eimeria and Isospora spp.20, 123, 176 The diagnosis of disease caused by E. coli is therefore made with certainty only after bacteriological examination.
Specimens for bacteriological examination should consist of short sections of affected intestinal tract, several mesenteric lymph nodes, and tissue blocks from organs such as the liver, spleen, kidneys and brain, preferably from animals that have not been treated with antimicrobial drugs. Tissue specimens must be collected using aseptic techniques, as soon after the death of the animal as possible, and must be placed in individual, sterile containers. In an outbreak of the disease, specimens from more than one animal should be taken. As E. coli may spread from the intestine after death, it is imperative to note time between death and the necropsy, as well as the amount of decomposition that has taken place.
Rectal swabs are less satisfactory as specimens for bacteriological examination as the faecal flora in the rectum may bear little relation to the situation in the small intestine. If intestinal samples are not available, rectal swabs from several untreated animals suffering from diarrhoea should be taken, if possible.
The samples should be cultured on a medium such as MacConkey agar, which allows differentiation of E. coli from other bacteria. A blood agar medium should also be inoculated in order to detect other possible bacterial causes of the disease and to identify haemolytic colonies. It is essential to select three to five smooth colonies per culture for serotyping, as non-pathogenic intestinal strains may be present together with pathogenic strains. Serotyping is usually carried out by slide agglutination and may be confirmed by tube agglutination tests. The pathogenic serotype is usually clearly predominant. If enteric colibacillosis is not the cause of the disease, a mixed growth of various serotypes in more-or-less equal numbers is found.
Tests for virulence factors are more definitive than is serotyping. However, several of the pili, notably K99, 987P, and F18, are not readily expressed on bacteria grown on routine culture media.37, 125 Special media have been developed for expression of K99 and F18 pili. The enterotoxins can be assayed by a number of biological methods present in only specialized laboratories. Immunological tests have been developed for detection of LT and STa.
The diagnosis of dysentery due to STEC is based on the clinical picture and the recovery from the faeces of STEC of a serotype known to be implicated in disease in calves. If dead calves are examined the classical AE lesions can be demonstrated in the terminal ileum and colon. The diagnosis of watery diarrhoea due to STEC is more challenging. In some cases, bloody diarrhoea may have been noticed in some calves in a herd while others had watery diarrhoea. Isolation of an STEC which produces Stx1 and possesss the eae gene is suggestive of STEC-mediated diarrhoea.
The diagnosis of oedema disease is based on the epidemiology, clinical signs, gross and microscopic pathology, and the isolation of haemolytic E. coli from the small intestine. 10, 88 However, the E. coli may have decreased in numbers or disappeared from the intestine by the time the pig is examined. Stx2e present in intestinal contents can be detected in Vero cell culture.
The detection of virulence genes is now a valuable method for identification of E. coli that are enteric pathogens. Gene probes are effective in identifying the presence of virulence genes,118 but polymerase chain reaction (PCR) amplification is a more rapid and simpler method of detecting virulence genes in E. coli isolated from animals with diarrhoea or oedema disease.50, 95, 102, 147 A single multiplex reaction may be used to detect genes for K99, F41, STa, Eae, Stx1, and Stx2.50
Septicaemic disease can be identified on the basis of culture of blood or organs of affected animals.
Differential diagnosis
The most commonly encountered diseases which may be confused with enteric colibacillosis are nutritional scours caused by ingestion of excessive quantities of milk by neonatal animals, and, depending on the animal species involved, infections caused by agents such as rotavirus, coronavirus, Campylobacter jejuni, Salmonella, Clostridium perfringens types A, B and C, Brachyspira hyodysenteriae, and Cryptosporidium, Eimeria and Isospora spp.20, 97 Peracute or acute oedema disease in pigs should be differentiated from mulberry heart disease (vitamin E/selenium deficiency) and the subacute or chronic disease manifested by neurological signs from conditions such as bacterial meningoencephalitis caused by Haemophilus parasuis or Streptococcus suis, haemagglutinating encephalomyelitis virus, and water deprivation (salt poisoning). Laboratory tests are required to rule out these possibilities.
Control
The factors that can influence the outcome of the host-parasite interaction should be examined and those that might contribute to disease should be corrected. The most common predisposing factors include the quality and quantity of colostrum taken in, climatic changes, mixing of young animals from different origins, and general hygiene. Cold can exacerbate or precipitate scours in pigs. An environmental temperature of 28 to 30 °C should be maintained for newly weaned pigs.
Vaccination of pregnant animals is effective in controlling E. coli diarrhoea in neonatal animals.124, 126, 128, 129 Pilus-based vaccines are popular because only a few types of pili need to be included to ensure protection against all the common serotypes of ETEC. Anti-pilus antibody has an anti-adhesive effect which prevents intestinal attachment and colonization by ETEC in cattle, sheep and pigs.64, 89, 90 Bovine vaccines should contain K99 pilus antigen, ovine vaccines F41 and K99, and porcine vaccines K88, K99 and 987P. Two vaccinations should be given to pregnant animals at a monthly interval, the second one preferably a month before parturition. Previously vaccinated pregnant animals need be given only a single vaccination about one month before parturition.
The two vaccines prepared by Onderstepoort Biological Products, one for cattle and sheep and the other for pigs, contain formalinized whole-cell bacterins from smooth strains emulsified in oil. The strains included are reviewed periodically, so that common southern African pathogenic strains of E. coli are always represented.75 The vaccine for cattle and sheep always contains K99 pili, and the one for pigs, K88 pili. Septicaemia-causing strains such as E. coli O78:K80 are included in the cattle and sheep vaccines, while the two most common South African serotypes occurring in pigs, E. coli O149:K91 and O141:K85, are incorporated into the porcine vaccine.
Specific antibody to K99 may be given orally to calves in the face of an outbreak of ETEC diarrhoea among calves from unvaccinated dams.86
Early intake of adequate amounts of colostrum by the new-born animals is critical for providing both specific antibodies and non-specific factors that contribute to protection against enteric and septicaemic diseases due to E. coli. The intake of colostrum should equal at least 10 per cent of a young animal’s body weight by 24 hours after birth, and the initial ingestion of colostrum should occur within the first few hours of life to maximize the absorption of antibodies. 53
Mismothering, teat defects and agalactia must be identified by the stockman, who must administer colostrum by bottle when the need arises. Colostrum banks, frozen or freeze-dried, should be kept for such emergencies. Some cows produce colostrum which has a low globulin content and their calves remain hypogammaglobulinaemic despite ingestion of the correct amount of colostrum.
There is no consensus on the methods that should be used to control post-weaning diarrhoea and oedema disease in piglets. Immunization of sows effectively allows for passive protection of nursing piglets, but antibodies need to be added to the feed or administered orally to protect weaned pigs.88 Weaner piglets can mount an immune response but their active response to ETEC may be suppressed by maternal antibodies. Stimulating active immunity by feeding inactivated ETEC during the suckling period is claimed to be effective against post-weaning infections, but it is not widely practised. Addition of acidifiers and zinc oxide to the feed are commonly used methods for reducing the problem of post-weaning diarrhoea due to E. coli.64 Probiotics have been evaluated in experimental infections but have not been effective.36
Animals suffering from diarrhoea should be treated to restore the fluid balance, as death usually results from dehydration. 20 Oral or parenteral electrolytes may be administered, depending on the severity of the disease. Oral antibiotic therapy is sometimes advocated to suppress the growth of ETEC, but variable and increasing antibiotic resistance is a major problem. An antibiogram should therefore be performed on the isolate causing an outbreak. Antimicrobials in feed or water should be administered for four to five consecutive days, bearing in mind that the appetite of affected animals may be depressed. Colisepticaemia should be treated with injectable antimicrobials.
As with post-weaning scours, no single recommendation can be made for the control of oedema disease. Restricting feed intake, controlling the ambient temperature, and use of antibiotics to reduce levels of the pathogenic E. coli may reduce the prevalence of the disease. A vaccine made from glutaraldehyde-inactivated Stx2e has proved effective in protecting pigs vaccinated prior to weaning.94 Passive immunization with antibodies against this toxoid also protected pigs from oedema disease.
A vaccine produced from a rough strain of E. coli has been effective in protecting cattle against E. coli mastitis.79, 80
References
- AARESTRUP, F.M., JORSAL, S.E., AHRENS, P., JENSEN, N.E. & MEYLING, A., 1997. Molecular characterization of Escherichia coli strains isolated from pigs with edema disease. Journal of Clinical Microbiology, 35, 20–24.
- ACRES, S.D., 1985. Enterotoxigenic Escherichia coli infections in newborn calves: A review. Journal of Dairy Science, 68, 229–256.
- ARBUCKLE, J.B.R., 1968. The occurrence of Escherichia coli somatic antibody in pig serum, colostrum and milk and an investigation of its possible significance in immunity. British Veterinary Journal, 124, 273–281.
- ARMSTRONG, W.D. & CLINE, T.R., 1977. Effects of various nutrient levels and environmental temperatures on the incidence of colibacillary diarrhoea in pigs: Intestinal fistulation and titration studies. Journal of Animal Science, 45, 1042–1050
- BAKER, D.R., BILLEY, L.O. & FRANCIS, D.H., 1997. Distribution of K88 Escherichia coli -adhesive and nonadhesive phenotypes among pigs of four breeds. Veterinary Microbiology, 54, 23–32.
- BANKS, K.L. & MCGUIRE, T.C., 1989. Neonatal immunology. In: HALLIWELL, R.E.W. & GORMAN, N.T., (eds) Veterinary Clinical Immunology. Philadelphia, London, Toronto, Montreal, Sydney, Tokyo: W.B. Saunders Company
- BERTIN, Y., MARTIN, C., GIRARDEAU, J.P., POHL, P. & CONTREPOIS, M., 1998. Association of genes encoding P fimbriae, CS31A antigen and EAST 1 toxin among CNF1-producing Escherichia coli strains from cattle with septicemia and diarrhea. FEMS Microbiol Letters 162, 235–239.
- BERTSCHINGER, H.U., BACHMANN, M., METTLER, C., POSPISCHIL, A., SCHRANER, E.M., STAMM, M., SYDLER, T. & WILD, P., 1990. Adhesive fimbriae produced in vivo by Escherichia coli 0139:K12(B):H1 associated with enterotoxaemia in pigs. Veterinary Microbiology, 25, 267–281.
- BERTSCHINGER, H.U., EGGENBERGER, E., JUCKER, H. & PFIRTER, H.P., 1979. Evaluation of low-nutrient, high-fibre diets for the prevention of porcine Escherichia coli enterotoxaemia. Veterinary Microbiology, 3, 281–290.
- BERTSCHINGER, H.U. & GYLES, C.L., 1994. Oedema Disease, In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans, Oxon, UK: CAB International.
- BERTSCHINGER, H.U. & NIELSEN, N.O., 1992. Edema disease. In: LEMAN, A.D., STRAW, B., MEGELING, W.L., D’ALLAIRE, S. & TAYLOR, D.J., (eds). Diseases of Swine. 7th edn. Ames, Iowa: Iowa State University Press
- BERTSCHINGER, H.U. & POHLENZ, J., 1983. Bacterial colonization and morphology of the intestine in porcine Escherichia coli enterotoxemia (edema disease). Veterinary Pathology, 20, 99–110.
- BERTSCHINGER, H.U., POHLENZ, J. & ROSS, R.F., 1986. Coliform mastitis. In: leman, a.d., straw, b., glock, r.d., mengeling, w.l., penny, r.h.c. & scholl, e., (eds). Diseases of Swine. 6th edn. Ames, Iowa: Iowa State University Press.
- BESSER, T.E. & GAY, C.C., 1985. Septicemic colibacillosis and failure of passive transfer of colostral immunoglobulin in calves. Veterinary Clinics of North America, Food Animal Practice, 1, 445–459.
- BESSER T.E., SZENCI, O. & GAY, C.C., 1990. Decreased colostral immunoglobulin absorption in calves with postnatal acidosis. Journal of the American Veterinary Medical Association, 196, 1239–1243.
- BETTELHEIM, K.A., 1994. Biochemical characteristics of Escherichia coli. In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans. Oxon, UK: CAB International.
- BEUTIN, L., MONTENEGRO, M.A., ΦRSKOV, I,. ΦRSKOV, F., PRADA, J., ZIMMERMAN, S. & STEPHAN, R., 1989. Close association of verotoxin (Shiga-like toxin) production with enterohemolysin production in strains of Escherichia coli. Journal of Clinical Microbiology, 27, 2559– 2564.
- BIJLSMA, I.G.W., DE NIJS, A., VAN DER MEER, C. & FRIK, J.F., 1982. Different pig phenotypes affect adherence of Escherichia coli to jejunal brush borders by K88ab, K88ac or K88ad antigen. Infection and Immunity, 37, 891–894.
- BINNS, M.M., DAVIES, D.L. & HARDY, K.G., 1979. Cloned fragments of the plasmid ColV, I-K94 specifying virulence and serum resistance. Nature (London), 279, 778–781.
- BLOOD, D.C. & RADOSTITS, O.M., 1989. Veterinary Medicine. 7th edn. London, Philadelphia, Sydney, Tokyo, Toronto: Baillière Tindall.
- BORCZYK, A.A., KARMALI, M.A., LIOR, H. & DUNCAN, L.M.C., 1987. Bovine reservoir for verotoxin-producing Escherichia coli O157:H7. Lancet, 1, 98.
- BORMAN-EBY, H.C., MCEWAN, S.A., CLARKE, R.C., MCNAB, W.B., RAHN, K. & VALDISIESO-GARCIA, A., 1993. The seroprevalence of verocytotoxin producing Escherichia coli in Ontario dairy cows and associations with production and management. Preventive Veterinary Medicine, 15, 261–274.
- BOYD, J.W., 1974. Neonatal diarrhoea in calves. The Veterinary Record, 95, 310–313.
- BROWN, J.E., ECHEVERRIA, P, & LINDBERG, A.A., 1991. Digalactosylcontaining glycolipids as cell-surface receptors for Shiga toxin of Shigella dysenteriae 1 and related cytotoxins of Escherichia coli. Reviews of Infectious Diseases, 13 (Suppl. 4), S298–303.
- CANTEY, J.R. & BLAKE, R.K., 1977. Diarrhoea due to Escherichia coli in the rabbit: A novel mechanism. Journal of Infectious Diseases, 135, 454–462.
- CHANTER, N., HALL, G.A., BLAND, A.P., HAYLE, A.J. & PARSONS, D.R., 1986. Dysentery in calves caused by an atypical strain of Escherichia coli (S102-9). Veterinary Microbiology, 12, 241–253.
- CHANTER, N., MORGAN, J.H., BRIDGER, J.C., HALL, G.A. & REYNOLDS, D.J., 1984. Dysentery in gnotobiotic calves caused by atypical Escherichia coli. The Veterinary Record, 114, 71.
- CHINA, B., PIRSON, V. & MAINIL, J., 1998. Prevalence and molecular typing of attaching and effacing Escherichia coli among calf populations in Belgium. Veterinary Microbiology, 63, 249–259
- CHINA, B., JACQUEMIN, E., DEVRIN, A.C., PIRSON, V. & MAINIL, J., 1999. Heterogeneity of the eae genes in attaching/effacing Escherichia coli from cattle: Comparison with human strains. Research in Microbiology, 150, 323–332.
- CLUGSTON, R.E., NIELSEN, N.O. & SMITH, D.L.T., 1974. Experimental edema disease of swine (E. coli enterotoxaemia). III. Pathology and pathogenesis. Canadian Journal of Comparative Medicine, 38, 34–43.
- CONWAY, P.L., WELIN, A. & COHEN, P.S., 1990. Presence of K88-specific receptors in porcine ileal mucus is age-dependent. Infection and Immunity, 58, 3178–3182.
- COOKE, E.M., 1985. Escherichia coli — an overview. Journal of Hygiene, Cambridge, 95, 523–530
- DEAN, E.A., 1990. Comparison of receptors for 987P pili of enterotoxigenic Escherichia coli in the small intestine of neonatal and older pigs. Infection and Immunity, 58, 4030–4035
- DEAN, A.G., CHING, Y.-C., WILLIAMS, R.G. & HARDEN, L.B., 1972. Test for Escherichia coli enterotoxin using infant mice. Application in a study of diarrhea in children in Honolulu. Journal of Infectious Diseases, 125, 407–411.
- DEAN-NYSTROM, E.A., BOSWORTH, B.T., CRAY, W.C. JR & MOON H.W., 1997. Pathogenicity of Escherichia coli O157:H7 in the intestines of neonatal calves. Infection and Immunity, 65, 1842–1848
- DE CUPERE, F., DEPREZ, P., DEMEULENAERE, D. & MUYLLE, E., 1992. Evaluation of the effect of 3 probiotics on experimental Escherichia coli enterotoxaemia in weaned pigs. Journal of Veterinary Medicine B, 39, 277–284.
- DE GRAAF, F.K. & GAASTRA, W., 1994. Fimbriae of enterotoxigenic Escherichia coli. In: KLEMM, P., (ed.). Fimbriae: Adhesion, Genetics, Biogenesis, and Vaccines, Boca Raton: CRC Press
- DEPREZ, P., VAN DEN HENDE, C., MUYLLE, E. & OYAERT, W., 1986. The influence of the administration of sow’s milk on the postweaning excretion of haemolytic Escherichia coli in the pig. Veterinary Research Communications, 10, 469–478.
- DE RYCKE, J., GUILLOT, J.F. & BOIVIN, R., 1987. Cytotoxins in non-enterotoxigenic strains of Escherichia coli isolated from faeces of diarrhoeic calves. Veterinary Microbiology, 15, 137–150.
- DE RYCKE, J., MILON, A. & OSWALD, E., 1999. Necrotoxic Escherichia coli (NTEC): Two emerging categories of human and animal pathogens. Veterinary Research, 30, 221–233.
- DOBRESCU, L., 1983. New biological effect of edema disease principle (Escherichia coli neurotoxin) and its use as an in vitro assay for this toxin. American Journal of Veterinary Research, 44, 31–34.
- DUBREUIL, J.D., 1997. Escherichia coli STb enterotoxin. Microbiology, 143, 1783–1795.
- DUHAMEL, G.E., MOXLEY, R.A., MADDOX, C.W. & ERICKSON, E.D., 1992. Enteric infection of a goat with enterohemorrhagic Escherichia coli (O103:H2). Journal of Veterinary Diagnostic Investigation, 4, 197–200.
- DUNNE, H.W., 1975. Colibacillosis and edema disease. In: DUNNE, H.W. & LEMAN, A.D, (eds). Diseases of Swine. 4th edn. Ames, Iowa: Iowa State University Press. 674.
- FAIRBROTHER, J.M., BROES, A., JACQUES, M. & LARIVIERE, S., 1989. Pathogenicity of Escherichia coli O115:K‘V165’ strains isolated from pigs with diarrhea. American Journal of Veterinary Research, 50, 1029–1036.
- FAIRBROTHER, J.M. & NGELEKA, M., 1994. Extraintestinal E. coli infections in pigs. In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans, Oxon, UK: CAB International.
- FERNANDEZ-BEROS, M.E., KISSEL, V., LIOR, H. & CABELLO, F.C., 1990. Virulence-related genes in ColV plasmids of Escherichia coli isolated from human blood and intestines. Journal of Clinical Microbiology, 28, 742–746.
- FEY, H., 1972. Colibacillosis in calves. Bern, Switzerland: Hans Huber.
- FRANCIS, D.H., GRANGE, P.A., ZEMAN, D.H., BAKER, D.R., SUN, R. & ERICKSON, A.K., 1998. Expression of mucin-type glycoprotein K88 receptors strongly correlates with piglet susceptibility to K88(+) enterotoxigenic Escherichia coli, but adhesion of this bacterium to brush borders does not. Infection and Immunity, 66, 4050–4055.
- FRANCK, S.M., BOSWORTH, B.T. & MOON, H.W., 1998. Multiplex PCR for enterotoxigenic, attaching and effacing, and Shiga toxin-producing Escherichia coli strains from calves. Journal of Clinical Microbiology, 36,1795–1797.
- GANNON, V.P.J., GYLES, C.L. & FRIENDSHIP, R.W., 1988. Characteristics of verotoxigenic Escherichia coli from pigs. Canadian Journal of Veterinary Research, 52, 331–337
- GAY, C.C., 1984. Failure of passive transfer of colostral immunoglobulin and neonatal disease in calves: A review. In: acres, s., (ed.). Proceedings of the Fourth International Symposium on Neonatal Diarrhea, Saskatoon, Saskatchewan, Veterinary Infectious Diseases Organisation
- GAY, C.C. & BESSER, T.E., 1994. Escherichia coli septicaemia in calves. In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans, Oxon, UK: CAB International.
- GAY, C.C., MCKAY, K.A. & BARNUM, D.A., 1964. Studies on colibacillosis of calves. I. The antibody acquired by calves as the result of vaccination of the dam. Canadian Veterinary Journal, 5, 248–261.
- GILL, D.M., CLEMENTS, J.D., ROBERTSON, D.C. & FINKELSTEIN, R.A., 1981. Subunit number and arrangement in Escherichia coli heat-labile enterotoxin. Infection and Immunity, 33, 677–682.
- GILL, D.M. & RICHARDSON, S.H., 1980. Adenosine diphosphateribosylation of adenylate cyclase catalyzed by heat-labile enterotoxin Escherichia coli: Comparison with cholera toxin. Journal of Infectious Diseases, 141, 64–70.
- GIRARDEAU, J.P., DER-VARTANIAN, M., OLLIER, J.L. & CONTREPOIS, M., 1988. CS31A, a new K88-related fimbrial antigen on bovine enterotoxigenic and septicemic Escherichia coli strains. Infection and Immunity, 56, 2180–2188.
- GONZALES, E.A. & BLANCO, J., 1985. Production of cytotoxin VT in enteropathogenic and non-enteropathogenic Escherichia coli strains of porcine origin. FEMS Microbiological Letters, 26, 127–130.
- GREEN, S.L. & SMITH, L.L., 1992. Meningitis in neonatal calves: 32 cases (1983–1990). Journal of the American Veterinary Medical Association, 201, 125–128.
- GYLES, C.L., 1992. Escherichia coli enterotoxins. In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans, Oxon, UK: CAB International.
- GYLES, C.L., PALCHAUDHURI, S. & MAAS, W.K., 1977. Naturally occurring plasmid carrying genes for enterotoxin production and drug resistance. Science, 198, 198–199.
- HALL, G.A., REYNOLDS, D.J., CHANTER, N., MORGAN, J.H., PARSONS, K.R., DEBNEY, T.G., BLAND, A.P. & BRIDGER, J.C., 1985. Dysentery caused by Escherichia coli (SlO2-9) in calves: Natural and experimental disease. Veterinary Pathology, 22, 156–163.
- HAMPSON, D.J., 1986. Alterations in piglet small intestine structure a weaning. Research in Veterinary Science, 40, 32–40.
- HAMPSON, D.J., 1994. Postweaning Escherichia coli Diarrhoea in Pigs. In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans. Oxon, UK: CAB International.
- HAMPSON, D.J., FU, Z.F., BETTLEHEIM, K.A. & WILSON, M.W., 1988. Managemental influences on the selective proliferation of two strains of haemolytic Escherichia coli in weaned pigs. Epidemiology and Infection, 100, 213–220
- HAMPSON, D.J., FU, Z.F. & ROBERTSON, I.D., 1987. Investigation of the source of haemolytic Escherichia coli infecting weaned pigs. Epidemiology and Infection, 99, 149–153.
- HAMPSON, D.J., FU, Z.F. & SMITH, W.C., 1988. Pre-weaning supplementary feed and porcine post weaning diarrhoea. Research in Veterinary Science, 44, 309–314.
- HAMPSON, D.J. & SMITH, W.C., 1986. Influence of creep feeding and dietary intake after weaning on malabsorption and occurrence of diarrhoea in the newly weaned pig. Research in Veterinary Science, 41, 63–69.
- HAMPSON, D.J., WOODWARD, J.M. & CONNAUGHTON, I.D., 1993. Genetic analysis of Escherichia coli from porcine postweaning diarrhoea. Epidemiology and Infection, 110, 575–581
- HARDIE, E.M. & KRUSE-ELLIOT, K., 1990. Endotoxic shock. Part 1: A review. Journal of Veterinary Internal Medicine, 4, 258–266.
- HAREL, J., LAPOINTE, H., FALLARA, A., LORFIE, L.A., BIGRAS-POULIN, M., LARIVIERE, S. & FAIRBROTHER, J.M., 1991. Detection of genes for fimbrial antigens and enterotoxins associated with Escherichia coli serogroups isolated from pigs with diarrhea. Journal of Clinical Microbiology, 29, 745–752.
- HARNETT, N.M. & GYLES, C.L., 1983. Enterotoxigenicity of bovine and porcine Escherichia coli of O groups 8, 9, 20, 64, 101, and X46. American Journal of Veterinary Research, 44, 1210–1214.
- HARNETT, N.M. & GYLES, C.L., 1985. Enterotoxin plasmids in bovine and porcine enterotoxigenic Escherichia coli of O groups 9, 20, 64 and 101. Canadian Journal of Comparative Medicine, 49, 79–87.
- HARNETT, N.M. & GYLES, C.L., 1985. Linkage of genes for heat-stable enterotoxin, drug resistance, K99 antigen, and colicin in bovine and porcine strains of enterotoxigenic Escherichia coli. American Journal of Veterinary Research, 46, 428–433.
- HENTON, M.M., 2002. Section of Bacteriology, Veterinary Research Institute, Onderstepoort. Unpublished data.
- HENTON, M.M. & ENGELBRECHT, M.M., 1997. Escherichia coli serotypes in pigs in South Africa. Onderstepoort Journal of Veterinary Research, 64, 175–187.
- HINTON, M., 1985. The sub-specific differentiation of Escherichia coli with particular reference to ecological studies in young animals including man. Journal of Hygiene, Cambridge, 95, 595–609.
- HIRSH, D.C., KIRKHAM, C. & WILSON, W.D., 1993. Characteristics of Escherichia coli isolated from septic foals. Veterinary Microbiology, 34, 123–130.
- HOGAN, J.S., SMITH, K.L., TODHUNTER, D.A. & SCHOENBERGER, P.S., 1992. Field trial to determine efficacy of an Escherichia coli J5 mastitis vaccine. Journal of Dairy Science, 75, 78–84.
- HOGAN, J.S., WEISS, W.P., TODHUNTER, D.A., SMITH, K.L. & SCHOENBERGER, P.S., 1992. Efficacy of an Escherichia coli J5 mastitis vaccine in an experimental challenge trial. Journal of Dairy Science, 75, 415–422
- HOLLAND, R.E., GRIMES, S.D., WALKER, R.D. & WILSON, R.A., 1996. Experimental inoculation of foals and pigs with an enterotoxigenic E. coli isolated from a foal. Veterinary Microbiology, 52, 249–257.
- HOLLAND, R.E., SCHMIDT, A., SRIRANGANATHAN, N., GRIMES, S.D., WILSON, R.A., BROWN, C.M. & WALKER, R.D., 1996. Characterization of Escherichia coli isolated from foals. Veterinary Microbiology, 48, 243–255
- HOLLAND, R.E., SRIRANGANATHAN, N. & DUPONT, L., 1989. Isolation of enterotoxigenic Escherichia coli from a foal with diarrhea. Journal of the American Veterinary Medical Association, 194, 389–391.
- HUNT, E., 1993. Diarrhoeal diseases of neonatal ruminants. In: HOWARD J.L., (ed.). Current Veterinary Therapy: Food Animal Practice, 3. Philadelphia: W.B. Saunders.
- IKE, K., KAMADA, M., ANZAI, T., IMAGAWA, H., KUMANOMIDO, T., NAKAZAWA, M., KASHIWAZAKI, M. & KUME, T., 1987. Some properties of Escherichia coli isolated from foals with diarrhoea and mares with metritis. Bulletin of the Equine Research Institute, 24, 33–41.
- IKEMORI, Y., KUROKI, M., PERALTA, R.C., YOKOYAMA, H. & KODAMA, Y., 1992. Protection of neonatal calves against fatal enteric colibacillosis by administration of egg yolk powder from hens immunized with K99-piliated enterotoxigenic Escherichia coli. American Journal of Veterinary Research, 53, 2005–2008.
- IMBERECHTS, H., BERTSCHINGER, H.U., STAMM, M., SYDLER, T., POHL, P., DE GREVE, H., HERNALSTEENS, J.P., VAN MONTAGU, M. & LINTERMANS P., 1994. Prevalence of F107 fimbriae on E. coli isolated from pigs with edema disease or post weaning diarrhoea. Veterinary Microbiology, 40, 219– 230.
- IMBERECHTS, H., DE GREVE, H. & LINTERMANS, P., 1992. The pathogenesis of edema disease in pigs. A review. Veterinary Microbiology, 31, 221–233.
- IMBERECHTS, H., DEPREZ, P., VAN DRIESSCHE, E. & POHL, P., 1997. Chicken egg yolk antibodies against F18ab fimbriae of Escherichia coli inhibit shedding of F18 positive E. coli by experimentally infected pigs. Veterinary Microbiology, 54: 328–341.
- ISAACSON, R.E., DEAN, E.A., MORGAN, R.L. & MOON, H.W., 1980. Immunization of suckling pigs against enterotoxigenic Escherichia coli-induced diarrhoeal disease by vaccinating dams with purified K99 or 987 P pili: Antibody production in response to vaccination. Infection and Immunity, 29, 824–826.
- ISAACSON, R.E., MOON, H.W. & SCHNEIDER, R.A., 1978. Distribution and virulence of Escherichia coli in the small intestines of calves with and without diarrhea. American Journal of Veterinary Research, 39, 1750– 1755
- JANKE, B.H., FRANCIS, D.H., COLLINS, J.E., LIBAL, M.C., ZEMAN, D.H. & JOHNSON, D.D., 1989. Attaching and effacing Escherichia coli infections in calves, pigs, lambs, and dogs. Journal of Veterinary Diagnostic Investigation, 1, 6–11.
- JANKE, B.H., FRANCIS, D.H., COLLINS, J.E., LIBAL, M.C., ZEMAN, D.H., JOHNSON, D.D. & NEIGER, R.D., 1990. Attaching and effacing Escherichia coli infection as a cause of diarrhoea in young calves. Journal of the American Veterinary Medical Association, 196, 897–901.
- JOHANSEN, M., ANDRESEN, L.O., JORSAL, S.E., THOMSEN, L.K., WADDELL, T.E. & GYLES, C.L., 1997. Prevention of edema disease in pigs by vaccination with verotoxin 2e toxin. Canadian Journal of Veterinary Research, 61, 280–285.
- JOHNSON, W.M., POLLARD, D.R., LIOR, H., TYLER, S.D. & ROZEE, K.R., 1990. Differentiation of genes coding for Escherichia coli verotoxin 2 and the verotoxin associated with porcine edema disease (VTe) by the polymerase chain reaction. Journal of Clinical Microbiology, 28, 2351– 2353
- JOHNSTON, N.E., ESTRELLA, R.A. & OXENDER, W.D., 1977. Resistance of neonatal calves given colostrum diet to oral challenge with a septicaemia-producing Escherichia coli. American Journal of Veterinary Research, 38, 1323–1326.
- JUBB, K.V.F., KENNEDY, P.C. & PALMER, N., 1985. Pathology of Domestic Animals. 3rd edn. New York: Academic Press.
- KARCH, H., SCHMIDT, H. & BRUNDER, W., 1998. Plasmid-encoded determinants of Escherichia coli O157:H7. In: KAPER, J.B. & O’BRIEN, A.D., (eds), Escherichia coli O157:H7 and Other Shiga Toxin-Producing E. coli Strains.Washington, D.C.: American Society for Microbiology.
- KARMALI, M.A., 1989. Infection by verocytotoxin-producing Escherichia coli. Clinical Microbiological Reviews, 2, 15–38.
- KAUSCHE, F.M., DEAN, E.A., ARP, L.H., SAMUEL, J.E. & MOON, H.W., 1992. An experimental model for subclinical edema disease (Escherichia coli enterotoxaemia) manifest as vascular necrosis in pigs. American Journal of Veterinary Research, 53, 281–287.
- KELLY, D., O’BRIEN, J.J. & MCCRACKEN, K.J., 1990. Effect of creep feeding on the incidence, duration and severity of post-weaning diarrhoea in pigs. Research in Veterinary Science, 49, 223–228
- KELLY, D., SMYTH, J.A. & MCCRACKEN, D.J., 1990. Effect of creep feeding on structural and functional changes of the gut of early weaned pigs. Research in Veterinary Science, 48, 350–356.
- KHAKHRIA, R., DUCK, D. & LIOR, H. 1990. Extended phage-typing scheme for Escherichia coli O157:H7. Epidemiology and Infection, 105, 511–520.
- KURTZ, H.J., BERGELAND, M.E. & BARNES, D.M., 1969. Pathologic changes in edema disease of swine. American Journal of Veterinary Research, 30, 791–806.
- LE BOUGUENEC, C. & BERTIN, Y., 1999. AFA and F17 adhesins produced by pathogenic Escherichia coli strains in domestic animals. Veterinary Research, 30, 317–342.
- LEVINE, M.M., 1987. Escherichia coli that cause diarrhoea: Enterotoxigenic, enteropathogenic, enteroinvasive, enterohaemorrhagic and enteroadherent. Journal of Infectious Diseases, 155, 377–389.
- LINGGOOD, M.A., ROBERTS, M., FORD, S., PARRY, S.H. & WILLIAMS, P.H., 1987. Incidence of the aerobactin iron uptake system among E. coli isolates from infection of farm animals. Journal of General Microbiology, 133, 835–842.
- LIOR, H., 1994. Classification of Escherichia coli. In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans, Oxon, UK: CAB International.
- LOGAN, E.F., 1974. Colostral immunity to colibacillosis in the neonatal calf. British Veterinary Journal, 130, 405–412
- LOGAN, E.F. & PENHALE, W.J., 1971. The prevention of experimental colisepticaemia by the intravenous administration of a bovine serum IgM-rich fraction. The Veterinary Record, 89, 663–667.
- LOPEZ-ALVAREZ, J. & GYLES, C.L., 1980. Occurrence of the Vir plasmid among animal and human strains of invasive Escherichia coli. American Journal of Veterinary Research, 41, 769–774.
- MACLEOD, D.L. & GYLES, C.L., 1991. Immunization of pigs with a purified Shiga-like toxin II variant toxoid. Veterinary Microbiology, 29, 309–318.
- MACLEOD, D.L., GYLES, C.L. & WILCOCK, B.P., 1991. Reproduction of edema disease of swine with purified Shiga-like toxin-II variant. Veterinary Pathology, 28, 66–73
- MCDANIEL, T.K., JARVIS, K.G., DONNENBERG, M.S. & KAPER, J.B., 1995. A genetic locus of enterocyte effacement conserved among diverse enterobacterial pathogens. Proceedings of the National Acadamy of Science, USA, 92, 1664–1668.
- MCDANIEL, T.K. & KAPER, J.B., 1997. A cloned pathogenicity island from enteropathogenic Escherichia coli confers the attaching and effacing phenotype on E. coli K-12. Molecular Microbiology, 23, 399–407.
- MAINIL, J., 1999. Shiga/verocytotoxins and Shiga/verotoxigenic Escherichia coli in animals. Veterinary Research, 30, 235–257.
- MAINIL, J.G., JACQUEMIN, E.R., KAECKENBEECK, A.E. & POHL, P.H., 1993. Association between the effacing (eae) gene and the Shiga-like toxin-encoding genes in Escherichia coli isolates from cattle. American Journal of Veterinary Research, 54, 1064–1068.
- MAINIL, J.G., MOSELEY, S.L., SCHNEIDER, R.S., SUTCH, K., CASEY, T.A. & MOON, H.W., 1986. Hybridization of bovine Escherichia coli isolates with gene probes for four enterotoxins (StaP, STaH, STb, LT) and one adhesion factor (K99). American Journal of Veterinary Research, 47, 1145–1148.
- MARQUES, L.R.M., PEIRIS, J.S.M., CRYZ, S.J. & O’BRIEN, A.D., 1987. Escherichia coli strains isolated from pigs with edema disease produce a variant of Shiga-like toxin II. Federation of European Microbiological Societies Microbiology Letters, 44, 33–38.
- METHIYAPUN, S., POHLENZ, J.F.L. & BERTSCHINGER, H.U., 1984. Ultrastructure of the intestinal mucosa in pigs experimentally inoculated with an edema disease-producing strain of Escherichia coli (O139:K12:H1). Veterinary Pathology, 21, 516–520.
- MILLER, B.G., NEWBY, T.J., STOKES, C.R. & BOURNE, F.J., 1984. Creep feeding and post-weaning diarrhoea in piglets. The Veterinary Record, 114, 296–297.
- MILLER, B.G., NEWBY, T.J., STOKES, C.R., HAMPSON, D.J. & BOURNE, F.J., 1983 The role of dietary antigen in the aetiology of post-weaning diarrhoea. Annales de Recherches Vétérinaires, 14, 487–492.
- MOON, H.W., 1969. Enteric colibacillosis in the newborn pig: Problems of diagnosis and control. Journal of the American Veterinary Medical Association, 155, 1853–1859.
- MOON, H.W., 1981. Protection against enteric colibacillosis in pigs suckling orally vaccinated dams: Evidence for pili as protective antigens. American Journal of Veterinary Research, 42, 173–177.
- MOON, H.W., 1990. Colonization factor antigens of enterotoxigenic Escherichia coli in animals. Current Topics in Microbiology and Immunology, 151, 147–165.
- MOON, H.W. & BUNN, T.O., 1993. Vaccines for preventing enterotoxigenic Escherichia coli infections in farm animals. Vaccine, 11, 213–220.
- MOON, H.W. & MCDONALD, J.S., 1983. Antibody response of cows to Escherichia coli pilus antigen K99 after oral vaccination with live or dead bacteria. American Journal of Veterinary Research, 44, 493–496.
- MOON, H.W., ROGERS, D.G. & ROSE, R., 1988. Effects of an orally administered live Escherichia coli pilus vaccine on duration of lacteal immunity to enterotoxigenic Escherichia coli in swine. American Journal of Veterinary Research, 49, 2068–2071.
- MOON, H.W. & RUNNELS, P.L., 1983. Trials with somatic (O) and capsular (K) polysaccharide antigens of enterotoxigenic Escherichia coli as protective antigens in vaccines for swine. Proceedings of the Fourth International Symposium on Neonatal Diarrhoea, University of Saskatchewan, 558–569.
- MOON, H.W. & RUNNELS, P.L., 1984. The K99 adherence system in cattle. In: BOEDEKER, E.C., (ed.). Attachment of Organisms to the Gut Mucosa. Vol. I. Boca Raton, Florida: CRC Press.
- MORRIS, J.A., THORNS, C.J., WELLS, G.A.H., SCHOTT, A.C. & SOJKA, W.J., 1983. The production of F41 fimbriae by piglet strains of enterotoxigenic Escherichia coli that lack K88, K99 and 987P fimbriae. Journal of General Microbiology, 129, 2753–2759.
- NABUURS, M.J., HOOGENDOORN, A. & VAN ZIJDERVELD-VAN BEMMEL, A., 1996. Effect of supplementary feeding during the sucking period on net absorption from the small intestine of weaned pigs. Research in Veterinary Science, 61, 72–77.
- NAGY, B., 1980. Vaccination of cows with a K99 extract to protect new-born calves against experimental enterotoxic colibacillosis. Infection and Immunity, 27, 21–24.
- NAGY, B., CASEY, T.A., WHIPP, S.C. & MOON, H.W., 1992. Susceptibility of porcine intestine to pilus-mediated adhesion by some isolates of piliated enterotoxigenic Escherichia coli increases with age. Infection and Immunity, 60, 1285–1294.
- NAGY, B. & FEKETE, P.Z., 1999. Enterotoxigenic Escherichia coli (ETEC) in farm animals. Veterinary Research, 30, 259–284.
- NAGY, B., WHIPP, S.C., IMBERECHTS, H., BERTSCHINGER, H.U., DEAN-NYSTROM, E.A., CASEY, T.A. & SALAJKA, E., 1997. Biological relationship between F18ab and F18ac fimbriae of enterotoxigenic and verotoxigenic Escherichia coli from weaned pigs with oedema disease or diarrhoea. Microbial Pathogenesis, 22, 1–11.
- NATARO, J.P. & KAPER, J.B., 1998. Diarrheagenic Escherichia coli. Clinical Microbiology Reviews, 11, 142–201.
- NGELEKA, M., HAREL, J., JACQUES, M. & FAIRBROTHER, J.M., 1992. Characterization of a polysaccharide capsular antigen of septicemic Escherichia coli O115:K ‘V165’ :F165 and evaluation of its role in pathogenicity. Infection and Immunity, 60, 5048–5056.
- NGELEKA, M., MARTINEAU-DOIZE, B. & FAIRBROTHER, J.M., 1994. Septicemia-inducing Escherichia coli O115:K‘V165’F165(1) resists killing by porcine polymorphonuclear leukocytes in vitro: Role of F165(1) fimbriae and K‘V165’ O-antigen capsule. Infection and Immunity, 62, 398–404.
- NIELSEN, N.O., 1986. Edema disease. In: LEMAN, A.D., STRAW, B., GLOCK, R.D., MENGELING, W.L., PENNY, R.H.C. & SCHOLL, E., (eds). Diseases of Swine. 6th edn. Ames, Iowa: Iowa State University Press.
- NOLLET, H., DEPREZ, P., VAN DRIESSCHE, E. & MUYLLE, E., 1999. Protection of just weaned pigs against infection with F18+ Escherichia coli by non-immune plasma powder. Veterinary Microbiology, 65, 37–45.
- O’BRIEN, A.D. & HOLMES, R.K., 1987. Shiga and Shiga-like toxins. Microbiological Reviews, 51, 206–220.
- ORDEN, J.A., RUIZ-SANTA-QUITERIA, J.A., CID, D., GARCIA, S., SANZ, R. & DE LA FUENTE, R., 1998. Verotoxin-producing Escherichia coli (VTEC) and eae-positive non-VTEC in 1–30-days-old diarrhoeic dairy calves. Veterinary Microbiology, 63, 239–248.
- ORDEN, J.A., RUIZ-SANTA-QUITERIA, J.A., CID, D., GARCIA, S. & DE LA FUENTE, R., 1999. Prevalence and characteristics of necrotoxigenic Escherichia coli (NTEC) strains isolated from diarrhoeic dairy calves. Veterinary Microbiology, 66, 265–273.
- ORSKOV, F., 1984. Genus 1. Escherichia. In: KRIEG, N.R. & HOLT, J.G., (eds). Bergey’s Manual of Systematic Bacteriology. Baltimore, London: Williams and Wilkins.
- ORSKOV, F. & ORSKOV, I., 1992. Escherichia coli serotyping and disease in man and animals. Canadian Jourmal of Microbiology 38, 699–704.
- OSWALD, E., SCHMIDT, H., MORABITO, S., KARCH, H., MARCHES, O. & CAPRIOLI, A., 2000. Typing of intimin genes in human and animal enterohemorrhagic and enteropathogenic Escherichia coli: characterization of a new intimin variant. Infection and Immunity, 68, 64–71.
- PEARSON, G.R., BAZELEY, K.J., JONES, J.R., GUNNING, R.F., GREEN, M.J., COOKSON, A. & WOODWARD, M.J., 1999. Attaching and effacing lesions in the large intestine of an eight-month-old heifer associated with Escherichia coli O26 infection in a group of animals with dysentery. The Veterinary Record, 145, 370–373.
- PEARSON, G.R. & LOGAN, E.F., 1979. The pathogenesis of enteric colibacillosis in neonatal unsuckled calves. The Veterinary Record, 105, 159–164.
- PEARSON, G.R., WATSON, C.A., HALL, G.A. & WRAY, C., 1989. Natural infections with an attaching and effacing Escherichia coli in the small and large intestines of a calf with diarrhoea. The Veterinary Record, 124, 297–299.
- PEETERS, J.E., 1994. Escherichia coli infections in rabbits, cats, dogs, goats and horses. In: GYLES, C.L., (ed.). Escherichia coli in Domestic Animals and Humans. Oxon, UK: CAB International.
- PORTER, P., NOAKES, D.E. & ALLEN, W.D., 1970. Secretory IgA and antibodies to Escherichia coli in porcine colostrum and milk and their significance in the alimentary tract of the young pig. Immunology, 245–257.
- RICHARDS, W.P.C. & FRASER, C.M. 1961. Coliform enteritis of weaned pigs. A description of the disease and its association with hemolytic Escherichia coli. Cornell Veterinarian, 51, 245–257
- RIPPINGER, P., BERTSCHINGER, H.U., IMBERECHTS, H., NAGY, B., SORG, I., STAMM, M., WILD, P. & WITTIG, W., 1995. Designations F18ab and F18ac for the related fimbrial types F107, 2134P and 8813 of Escherichia coli isolated from porcine postweaning diarrhea and from edema disease. Veterinary Microbiology, 45, 281–295.
- ROUSSET, E. & DUBREUIL, J.D., 1999. Evidence that Escherichia coli STb enterotoxin binds to lipidic components extracted from the pig jejunal mucosa. Toxicon, 37,1529–1537.
- SARIDAKIS, H.O., EL GARED, S.A., VIDOTTO, M.C. & GUTH, B.E., 1997. Virulence properties of Escherichia coli strains belonging to enteropathogenic (EPEC) serogroups isolated from calves with diarrhea. Veterinary Microbiology, 54, 145–153.
- SARMIENTO, J.I., RUNNELS, P.L. & MOON, H.W., 1990. Effects of preweaning exposure to a starter diet on enterotoxigenic Escherichia coli-induced postweaning diarrhea in swine. American Journal of Veterinary Research, 51, 1180–1183.
- SCALETSKY, I.C., SILVA, M.L & TRABULSI, L.R., 1984. Distinctive patterns of adherence of enteropathogenic Escherichia coli to HeLa cells. Infection and Immunity, 45, 534–536.
- SCHULMAN, A., 1973. Effects of weaning on the pH changes of the contents of the piglet stomach and duodenum. Nordisk Veterinaermedicin, 25, 220–225.
- SEIMIYA, Y., OHSHIMA, K., ITOH, H., OGASAWARA, N., OKUTOMO, M. & TANAKA, S., 1992. Central nervous system lesions due to Escherichia coli infection in neonatal calves. Journal of Veterinary Medical Science, 54, 767–768.
- SELLWOOD, R., 1984. The K88 adherence system in swine. In: BOEDEKER, E.C., (ed.). Attachment of Organisms to the Gut Mucosa. Vol. I. Boca Raton, Florida: CRC Press.
- SHERMAN, D.M., ACRES, S.D., SADOWSKI, P.L., SPRINGER, J.A., BRAY, B. RAYBOULD, T.J.G. & MUSCOPLAT, C.C., 1983. Protection of calves against fatal enteric colibacillosis by orally administering E. coli K99-specific monoclonal antibody. Infection and Immunity, 42, 653–658.
- SHANKS, P.L., 1938. An unusual condition affecting the digestive organs of the pig. The Veterinary Record, 12, 356–358.
- SMITH, H.W., 1974. A search for transmissible pathogenic characters in invasive strains of Escherichia coli: The discovery of a plasmidcontrolled toxin and a plasmid-controlled lethal character closely associated or identical with colicine V. Journal of General Microbiology, 83, 95–111.
- SMITH, H.W. & GYLES, C.L., 1970. The relationship between two apparently different enterotoxins produced by enteropathogenic strains of Escherichia coli of porcine origin. Journal of Medical Microbiology, 3, 387–401.
- SMITH, H.W. & HALLS, S., 1967. Observations by the ligated intestinal segment and oral inoculation methods on Escherichia coli infections in pigs, calves, lambs and rabbits. Journal of Pathology and Bacteriology, 93, 499–529.
- SMITH, H.W. & HALLS, S., 1968. The production of oedema disease and diarrhoea in weaned pigs by the oral administration of Escherichia coli: Factors that influence the course of the experimental disease. Journal of Medical Microbiology, 1, 45–59.
- SMITH, H.W. & JONES, J.E.T., 1963. Observations on the alimentary tract and its bacterial flora in healthy and diseased pigs. Journal of Pathology and Bacteriology, 86, 387–412.
- SMITH, H.W. & HUGGINS, M.B., 1978. The influence of plasmid-determined and other characteristics of enteropathogenic Escherichia coli on their ability to proliferate in the alimentary tracts of piglets, calves and lambs. Journal of Medical Microbiology, 11, 471–492.
- SMITH, H.W. & LINGGOOD, M.A., 1971. Observations on the pathogenic properties of the K88, Hly and Ent plasmids of Escherichia coli with particular reference to porcine diarrhoea. Journal of Medical Microbiology, 4, 467–485.
- SNODGRASS, D.R., SMITH, M.L. & KRAUTIL, F.L., 1982. Interaction of rotavirus and enterotoxigenic Escherichia coli in conventionally-reared dairy calves. Veterinary Microbiology, 7, 51–60.
- SOJKA, W.J., 1965. Escherichia coli in Domestic Animals and Poultry. Review Series No. 7 of the Commonwealth Bureau of Animal Health, Weybridge.
- SUGAI, M., HATAZAKI, K., MOGAMI, A., OHTA, H., PERES, S.Y., HERAULT, F., HORIGUCHI, Y., MASUDA, M., UENO, Y., KOMATSUZAWA, H., SUGINAKA, H. & OSWALD, E., 1999. Cytotoxic necrotizing factor type 2 produced by pathogenic Escherichia coli deamidates a gln residue in the conserved G-3 domain of the rho family and preferentially inhibits the GTPase activity of RhoA and rac1. Infection and Immunity, 67, 6550–6557.
- TIZARD, I., 1987. Immunity in the foetus and newborn. In: TIZARD, I., (ed.). Veterinary Immunology. An Introduction. 3rd edn. Philadelphia, London, Toronto, Sydney, Tokyo, Hong Kong: W.B. Saunders Company.
- TO, S.C.M., 1984. F41 antigen among porcine enterotoxigenc Escherichia coli strains lacking K88, K99, and 987P pili. Infection and Immunity, 43, 549–554.
- TZIPORI, S., 1981. The aetiology and diagnosis of calf diarrhoea. The Veterinary Record, 108, 510–514.
- VAN BEERS-SCHREURS, H.M.G., VELLENGA, L., WENSING, T.H. & BREUKINK, H.J., 1992. The pathogenesis of the post-weaning syndrome in weaned piglets; a review. Veterinary Quarterly, 14, 29–34.
- VOGELI, P., MEIJERINK, E., FRIES, R., NEUENSCHWANDER, S., VORLANDER, N., STRANZINGER, G. & BERTSCHINGER, H.U., 1997. A molecular test for the detection of E. coli F18 receptors: A breakthrough in the struggle against edema disease and post-weaning diarrhea in swine. Schweizer Archiv für Tierheilkunde, 139, 479–484.
- WADDELL, T.E., LINGWOOD, C.A. & GYLES, C.L., 1996. Interaction of verotoxin 2e with pig intestine. Infection and Immunity, 64, 1714–1719.
- WATERS, V.L. & CROSA, J.H., 1991. Colicin V virulence plasmids. Microbiological Reviews, 55, 437–450.
- WIELER, L.H., BAUERFEIND, R. & BALJER, G., 1992. Characterization of Shiga-like toxin producing Escherichia coli (SLTEC) isolated from calves with and without diarrhoea. Zentralblatt für Bakteriologie, 276, 243–253.
- WIELER, L.H., SCHWANITZ, A., VIELER, E., BUSSE, B., STEINBRUCK, H., KAPER, J.B. & BALJER, G., 1998. Virulence properties of Shiga toxin-producing Escherichia coli (STEC) strains of serogroup O118, a major group of STEC pathogens in calves. Journal of Clinical Microbiology, 36, 1604– 1607.
- WILSON, M.R., 1986. Enteric colibacillosis. In: LEMAN, A.D., STRAW, B., GLOCK, R.D., MENGELING, W.L., PENNY, R.H.C. & SCHOLL, E., (eds). Diseases of Swine. 6th edn. Ames, Iowa: Iowa State University Press. 520–528.
- WITTIG, W., KLIE. H., GALLIEN, P., LEHMANN, S., TIMM, M. & TSCHAPE, H., 1995. Prevalence of the fimbrial antigens F18 and K88 and of enterotoxins and verotoxins among Escherichia coli isolated from weaned pigs. Zentralblatt für Bakteriologie, 283, 95–104.
- WITTIG, W., PRAGER, R., STAMM, M., STRECKEL, W. & TSCHAPE, D.H., 1994. Expression and plasmid transfer of genes coding for the fimbrial antigen in porcine Escherichia coli strains. Zentralblatt für Bakteriologie, 281, 130–139.
- WOODS, P.R. & ROUSSEL, A.J., 1993. Colostrum and passive immunity. In: HOWARD, J.L., (ed.). Current Veterinary Therapy 3: Food Animal Practice. Philadelphia: W.B. Saunders Company.
- WOODWARD, J.M., CONNAUGHTON, I.D., FAHY, V.A., LYMBERY, A.J. & HAMPSON, D.J., 1993. Clonal analysis of Escherichia coli of serogroups 09, 020, and 0101 isolated from Australian pigs with neonatal diarrhoea. Journal of Clinical Microbiology, 31, 1185–1188.
- WRAY, C., MCLAREN, I. & PEARSON, G.R., 1989. Occurrence of ‘attaching and effacing’ lesions in the small intestine of calves experimentally infected with bovine isolates of verocytoxic E. coli. The Veterinary Record, 125, 365–368.
- WRAY, C. & MORRIS, J.A., 1985. Aspects of colibacillosis in farm animals. Journal of Hygiene, Cambridge, 95, 577–593.
- YAMAMOTO, T. & NAKAZAWA, M., 1997. Detection and sequences of the enteroaggregative Escherichia coli heat-stable enterotoxin 1 gene in enterotoxigenic E. coli strains isolated from piglets and calves with diarrhea. Journal of Clinical Microbiology, 35, 223–227.
- ZHU, C., HAREL, J., JACQUES, M., DESAUTELS, C., DONNENBERG, M.S., BEAUDRY, M. & FAIRBROTHER, J.M., 1994. Virulence properties and attaching-effacing activity of Escherichia coli O45 from swine postweaning diarrhea. Infection and Immunity, 62, 4153–4159.




