- Infectious Diseases of Livestock
- Part 3
- Histophilus somni disease complex in cattle
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Histophilus somni disease complex in cattle
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Histophilus somni disease complex in cattle
Synonym: Histophilosis
Previous author: P N THOMPSON
Current authors:
J D, TAYLOR,R
Professor, DVM, MPH, PhD, DACVIM (LA), DACVPM, Veterinary Pathobiology, College of Veterinary Medicine, Oklahoma State University, United States
TJ INZANA
Professor and Associate Dean for Research, College of Veterinary Medicine, Long Island University, 720 Northern Blvd., 216 Roth Hall, Brookville, NY 11548, USA
Introduction
Histophilus somni is the cause of a variety of clinical syndromes in cattle, including those associated with septicaemia, thrombotic meningoencephalitis, pneumonia, myocarditis, arthritis, mastitis, and disease of the reproductive tract in both cows and bulls. Various diseases due to H. somni can occur worldwide, although the incidence can vary greatly between countries and even within regions of the same country.
Disease due to H. somni was first described in 1956 in feedlots in the USA.53, 89 This disease manifested in the form of an infectious meningoencephalitis, and has been referred to as ‘sleeper syndrome’ (Figure 1), and more accurately, thrombotic meningoencephalitis (TME).59 Although TME was thought to be the major disease problem, fewer cases of it are being encountered, while respiratory disease and myocarditis are now recognized as the major diseases associated with H. somni.123 Histophilus somni is recognized as one of the four major bacterial agents of bovine respiratory disease (BRD) (Figure 2).
When disease due to H. somni first occurs in a region, it may present predominantly as TME, but as the infection becomes endemic there is a gradual change in its manifestation, and the respiratory form attains a higher profile.104
Histophilus somni is part of the normal mucosal flora of only ruminants, and can be readily isolated from healthy animals. Clinical disease is the result of prior infection (such as by viruses), or stress, due to crowding, shipping, lack of food or water, or environmental extremes.154, 155
There are several excellent reviews on H. somni, including the virulence factors and disease syndromes that it produces.24, 33, 59, 75, 84, 97, 134, 144, 149


Aetiology
The genus Histophilus consists of only H. somni, and belongs to the family Pasteurellaceae. Histophilus somni was formerly a member of the genus Haemophilus (blood loving) due to its requirement for blood or blood factors in the medium for growth. However, it was moved to the genus Histophilus based on molecular discrimination. Nonetheless, like haemophili of humans that are normal mucosal inhabitants in humans, H. somni is an obligate inhabitant of the mucous membranes of ruminants (sheep, cattle, bison, etc.). Histophilus somni is a small, pleomorphic, non-motile, Gram-negative, non-acid-fast coccobacillus that is a facultative anaerobe and requires CO2 for in vitro growth (capnophilic).75, 90
The prior species Haemophilus agni (isolated from sheep in North America) and Histophilus ovis (causes a form of meningoencephalitis in sheep similar to TME in cattle.127) are now recognized as belonging to a single taxon with H. somni.5, 148 Although there may be distinct biochemical differences164 and distinctions made by polymerase chain reaction (PCR)-based DNA fingerprinting and plasmid profile analysis,9 genetic similarity based on 16S rRNA and rpoB sequencing confirm that all isolates from these three former species are one species.5 Numerous multiplex PCR-based assays have been developed to identify H. somni and other BRD pathogens in clinical specimens.6, 15, 107, 178
There are substantial differences in virulence among H. somni isolates. In particular many isolates from the bovine prepuce are serum-sensitive and lack the virulence factors of BRD isolates.20, 28, 55, 75, 98, 143, 162 One bovine preputial isolate (strain 129Pt) whose genome has been completely sequenced20 lacks many of the virulence factors of BRD strain 2336.143 The virulence factors of H. somni (described below) protect the bacterium from host defences. Therefore, virulent isolates may be present as commensals and cause disease if they bypass innate host defences and gain access to systemic sites, whereas avirulent isolates appear to only live in the mucosal surfaces.30
An atmosphere containing 5 to 20 per cent carbon dioxide is required for primary isolation of H. somni. It grows well at 37 °C on media supplemented with 10 per cent blood or serum from cattle or sheep. However, the bacteria will also grow well in brain heart infusion or Columbia broth or agar supplemented with 0.1 per cent Trizma® base and 0.01 per cent thiamine monophosphate. Yeast extract (0.5 per cent) can also be added to further enhance growth of some strains.77 Histophilus somni is easily overgrown by most contaminants and a selective medium incorporating vancomycin, neomycin, nystatin, and cyclohexamide in 5 per cent horse blood agar can be used for isolation from clinical specimens that are not normally sterile.145
Histophilus somni is both X-factor (protoporphyrin, protohaeme, or haemin) and V-factor (nicotinamide adenine dinucleotide or NAD) independent, whereas senso stricto Haemophilus spp. require heat-stable X-factor and/or heat-labile V-factor for in vitro growth.141 The colonies are about 1 to 2 mm in diameter after two days’ growth and are non-haemolytic or weakly haemolytic (haemolysis may be enhanced by storage of culture plates at 4°C for several days), convex, circular, and pale yellow in colour. The colour of the colonies is more conspicuous when they are raised on a bacteriological loop or a white swab.177 Identification of H. somni is based on morphological and colony characteristics, independence of either X- or V-factors for growth, and biochemical testing.75 However, presumptive identification of H. somni as a disease agent can be made based on isolation of the bacterium from a systemic site of a specific host, requirement for blood and CO2 for growth, and colonies that are of yellow colour when picked up with a white swab. However, some isolates may be more tan in appearance. Respiratory and genital isolates of H. somni can be differentiated by biotyping (sugar fermentation) and a variety of PCR methods.10, 15, 42, 107, 178
Epidemiology
Several studies have indicated that subclinical carriers of H. somni probably play an important role in the epidemiology of the disease syndromes attributed to the organism, as they act as potential reservoirs of infection. Histophilus somni is maintained in the reproductive and respiratory mucosal tracts, which is supported by the fact that it is isolated from these sites in healthy cattle with relative frequency.30, 73, 74, 98, 139 In North America, the organism has also been isolated from the tonsils of commercially reared American bison (Bison bison).173 An evaluation of carriage of BRD pathogens in healthy cattle in the UK using qPCR indicated that 75 per cent of cattle carried H. somni in nasal passages, higher than Mannheimia haemolytica (13 per cent), but lower than Pasteurella multocida (95 per cent). Colonization rates and carriage of each of these species varied considerably over time.161 However, H. somni appears to be a more common commensal of the reproductive tract than the upper respiratory tract of healthy animals,30 and it can be isolated from the prepuce of a much higher percentage of bulls than it can from the vagina of cows.30, 73, 74, 75, 104 In one study of healthy cattle, cultures made from the prepuce, vagina and nasal cavity showed that 61 per cent, 15 per cent and 1.2 per cent, respectively, were positive for H. somni.30 Similar results were obtained in an abattoir survey of the urogenital tract of normal bulls; in addition, H. somni was recovered from the bladder, accessory sex glands and ampullae of about 20 per cent of the animals.104 However, many H. somni isolates from the bovine prepuce are serum-sensitive and avirulent,28 and therefore an unlikely source for disease transmission. In contrast, some strains isolated from the genital tract of apparently healthy cows caused fatal suppurative meningoencephalitis when inoculated intracisternally into the cisterna magna in calves, while others were nonpathogenic.98 In an endemic situation 40 to 60 per cent of cows in an asymptomatic herd may carry H. somni in the genital tract, and calves are colonized soon after birth. By the time they are seven months of age they are also carriers of H. somni in their genital tracts.104 Although disease is usually sporadic, up to 50 per cent or more of healthy animals may be seropositive.3, 36, 59, 68, 104
There is little documentation regarding the role of fomites in the spread of infection. Histophilus somni can survive for more than 70 days in secretions and discharges at freezing temperatures. At mild summer temperatures H. somni can survive for more than 70 days when mixed with blood and nasal mucus, but for less than 15 minutes in urine.36, 104 Despite this latter short survival period, it may not be sufficient to impede transmission by inhalation of urinary aerosols in cattle kept in close contact with each other.104 The respiratory system seems to be the most likely route of infection in natural cases of the disease,32, 59 as animals become infected by inhaling aerosols that contain the organism. In addition to inhalation, other possible means of transmission from the urogenital tracts of infected animals are by ingestion (licking) and venereally.75
A survey of a feedlot in Saskatchewan determined that histophilosis (considered a possible/likely agent in all possible disease manifestations) contributed to as much as 43 per cent of mortality in calves.168 These manifestations of fatal H. somni infection include myocarditis and pleuritis, occasionally TME, peracute septicaemia, and pneumonia. There was also circumstantial evidence suggesting that H. somni may be a pathogen in polyarthritis. Death from pneumonia mainly occurred during the first five weeks in the feedlot. Death from myocarditis, pleuritis, TME, septicaemia, and euthanasia because of polyarthritis occurred mainly after the third week in the feedlot.
The presence of H. somni in the upper respiratory tract (URT) can be relatively common, even in clinically healthy cattle. However, prevalence can vary by source farm, and may increase following transportation21 and presumably other stresses. As with other pathogens associated with BRD, it is unclear what stimulates proliferation and/or transmission of H. somni during these periods, although it is clearly a result of complex host-pathogen interactions. Similar questions remain regarding translocation from the URT to the lower respiratory tract (LRT), as well as its access to the vascular system and induction of septicaemia.123 The clinical diseases resulting from these processes have both unifying and unique themes.
The respiratory syndrome (i.e., BRD) is most common in calves under four months of age but also occurs in feedlot and adult cattle.30, 140 Histophilus somni is one of the most common bacteria isolated from fatal cases of calf pneumonia in Denmark.159 Calves are most susceptible when passive immunity wanes, with respiratory disease being most prevalent in spring and summer.140 Because H. somni is easily overgrown in vitro by other organisms, such as Mannheimia haemolytica, it is possible that the prevalence of respiratory disease due to H. somni may be underestimated.158
A recent study of the lower respiratory tract microbiome and resistome in cattle that died of BRD showed that H. somni, as well as M. haemolytica and Mycoplasma bovis were present more than 5 per cent of the time. Integrated and conjugative elements that harboured antimicrobial resistance genes were also relatively common, indicating the potential for antimicrobial drug failures and the need for susceptibility testing of isolates.93
A recent study of the occurrence of BRD pathogens in beef cattle in southern Brazil during a respiratory disease outbreak found that H. somni was the most common isolate from nasopharyngeal swabs (39 per cent), followed by bovine respiratory syncytial virus (35 per cent), bovine coronavirus (22 per cent), and M. haemolytica (13 per cent). Histophilus somni and M. haemolytica were identified from the lungs of at least one steer with fibrinous bronchopneumonia by PCR.64 Histophilus somni infection likely contributes to undifferentiated BRD of feedlot cattle,59, 75, 140, 142 and it may be isolated together with M. haemolytica18, 58, 61 or P. multocida185 pneumonic lesions both in unweaned calves and in older feedlot calves, and during experimental challenges. In such cases the bacteria are likely present as a poly-microbial biofilm.83 A study involving Canadian feedlots found that most calves had been exposed to H. somni prior to arrival at the feedlot.110 Calves with high antibody titres to H. somni on arrival had a reduced risk of developing respiratory disease. Another Canadian study showed that calves with higher H. somni antibody titres on arrival and increases in H. somni antibody titre after arrival had a decreased risk of developing undifferentiated fever.16
Thrombotic meningoencephalitis has been recorded in North and South America, Europe, Russia, Japan, New Zealand, South Africa and Zimbabwe.53, 60, 75, 92, 104, 163 The organism has been isolated from the respiratory and reproductive tracts of cattle in North America, South America, Europe,75 and likely anywhere that cattle are farmed.
Cattle between one and three years of age that have been in a feedlot for four weeks or longer are more likely to develop TME.12, 92, 140 However, the disease has also been diagnosed in calves less than four months of age, in adult dairy animals140 and in cattle on pasture.126, 163 It is most prevalent during the winter months,92, 105 and may be associated with recent transportation;105, 126 the mixing of cattle from different sources may also increase the risk of outbreaks. 140, 166 Ueno et al. classified 200 isolates of H. somni from Japan and other countries into nine major outer membrane protein (MOMP) clades (based on gene sequence) and 10 clusters based on pulsed field gel electrophoresis (PFGE) of DNA.166 Of interest is that TME isolates were significantly associated with MOMP clade 1a and PFGE cluster HS1. Following introduction of a bacterin vaccine using an HS1 strain of MOMP clade 1a, the incidence of TME due to this clonal group decreased, but the incidence of disease for other clonal groups increased.166
Histophilus somni-associated TME can be an important cause of mortality in feedlots.75 A survey of feedlot cattle in Ontario, Canada indicated that the population mortality rate due to TME was about 0,1 per cent of all the feedlot animals, while 12 per cent of all mortality was due to this condition.75 Bacteraemia and septicaemia are important preludes to infections resulting in mortality. In Great Britain, the incidence of septicaemia is reported to be increasing in calves between 6 and 20 months of age.21 Bacteraemia leads to localization in the brain (TME) and the heart (myocarditis), with more chronic cases resulting in abscesses in the left ventricular papillary muscle. Infection and necrosis of the laryngeal muscles may also occur.8, 123
Abortion in cattle as a result of H. somni infection has been reported in several countries.30, 59, 151 Few studies have examined disease syndromes in sheep in detail. However, an investigation of such syndromes in eight sheep from five farms in southern Brazil found animals afflicted with TME (4), septicaemia (4), bronchopneumonia (4), and myocarditis (3).64 Therefore, clinical manifestations of histophilosis in sheep appear to be similar to those in cattle.
Pathogenesis
A wide variation in the virulence of various H. somni isolates has been shown to exist through in vivo and in vitro experiments.55, 84, 98, 104 and supported by genome sequencing20, 143 Isolates from the prepuce may be non-pathogenic or produce mild lesions, while those from the brain may cause severe lesions using the proper experimental challenge, but little evidence of pneumonia; similarly those from the lung may cause severe pneumonia, but few lesions in the brain.55, 104 However, the same isolate can often be recovered from the brain and the lungs.
Some strains isolated from the female genital tract of apparently healthy cows caused severe fatal suppurative meningoencephalitis when inoculated intracisternally in calves, while others were non-pathogenic.98 It is possible that virulence is lost in subcultured organisms due to phase variation,104, 149 but this would not affect virulence in vivo.80 The complete genome sequence of a highly virulent pneumonia isolate (strain 2336)143 and an avirulent preputial isolate (strain 129Pt)20 has revealed that many of the genes responsible for virulence factors in the pneumonia isolate are lost or are pseudo-genes due to exogenous insertions and rearrangements in preputial strain 129Pt.
The major virulence factors of H. somni have been well established and include the heat-stable lipooligosaccharide (LOS), the high molecular weight immunoglobulin binding proteins (IbpA), survival within phagocytic cells, and biofilm formation. The LOS is an extremely important and versatile virulence factor, which occurs in the outer membrane,82 is a mediator of endothelial cell adhesion,23 can induce endothelial apoptosis,152 induces inflammation due its endotoxic activity82 and undergoes phase variation of the heterogeneous oligosaccharide to avoid the host immune response.80, 81 The galactose residues of the LOS oligosaccharide are decorated with terminal sialic acid in vivo,71, 78 which further nullifies the host immune response to the bacterium through inhibition of TLR4 signalling, while still accommodating phase variation of terminal galactose residues. Furthermore, sialylation of the LOS enhances binding of complement factor H, thereby enhancing resistance to serum- and polymorphonuclear leukocyte-mediated killing.76 At least one non-pathogenic preputial isolate (strain 129Pt) is unable to phase vary or sialylate its LOS oligosaccharide.78, 112 In addition to sialic acid, the H. somni LOS of virulent strains is decorated with phase variable phosphorylcholine (ChoP).70 Phosphorylcholine expression by H. somni results in aggregation of bovine platelets,96 and enhances colonization of the bovine upper respiratory tract, probably through binding to platelet activating factor on epithelial cells. However, ChoP also binds to C-reactive protein, which activates complement and enhances serum killing; therefore, phase variable ChoP is not expressed by H. somni in systemic sites.40 The structure of H. somni strain 738 LOS is shown in Figure 3.
Immunoglobulin binding protein is a large (120-350 kDa) protein on the cell surface in the form of a fibrillar network.27 High molecular weight IbpA has been found in virulent strains, but not in serum-sensitive strains from asymptomatic carriers, is shed from the bacterial cell surface and is associated with serum resistance and resistance to phagocytosis.27 IbpA is encoded by ibpA 153 and contains a duplicated 1.2-kb region (DR1 and DR2) at the C-terminal end. The repeats contain a Fic motif (HPFxxGNR) that is cytotoxic for epithelial, endothelial, and phagocytic cells.179, 183 Antibodies to this motif neutralize cytotoxic activity,183, 184 and immunization of mice and calves with a recombinant form of DR1 is protective against septicaemia and pneumonia in mouse and bovine models.43, 44 An open reading frame upstream of ibpA encodes for a protein (IbpB) with homology to proteins responsible for transport of large fibrillar proteins of other gram-negative bacteria, such as HMW1 and HMW2 in Haemophilus influenzae.182 Therefore, IbpA and IbpB likely function as a two-partner secretion system. The N-terminus region has a carbohydrate binding domain, which may contribute to cell adherence;153 mutations in this region also diminish biofilm formation. 136 For additional details and review of other H. somni outer membrane proteins the reader is referred elsewhere.26
Virulent, but not avirulent, strains of H. somni have long been shown to be capable of surviving within phagocytic cells,32, 49, 69, 72 which would make the organism a persistent intracellular pathogen. Intracellular survival has now been shown to be due to the ability of only virulent strains to block phagosome-lysosome fusion. Although a potential contributor, intracellular survival is not solely due to toxicity by the IbpA Fic motif in DR1 and DR2 because mutants lacking only this region become more serum susceptible, but intracellular survival is not affected. 125 Nonetheless, cell retraction due to collapse of the actin cytoskeleton is caused by the Fic motifs,179 resulting in cell retraction and transmigration of H. somni across the basement membranes.183
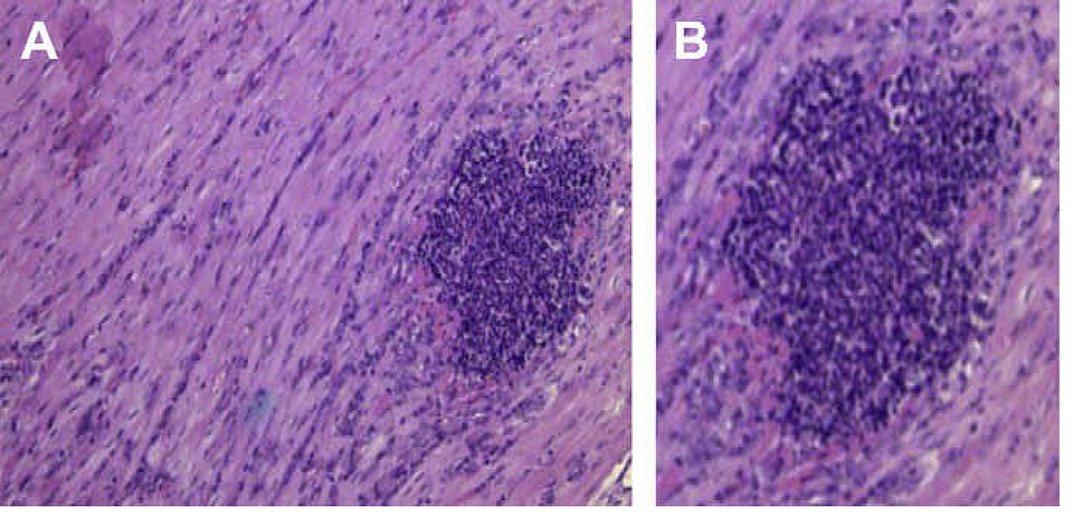
Histophilus somni (particularly virulent strains) is a prominent biofilm former (Figure 4), and its natural state is as a biofilm.137 During chronic infection (days or longer) a prominent biofilm can form in the myocardium (Figure 5 and Figure 6) and respiratory tract,135 and possibly other sites. Sudden death in calves has been attributed primarily to myocardial disease, but a biofilm infection has likely been present at that site for some time. Histophilus somni does not form a capsule, but only during biofilm formation a galacto-mannan exopolysaccharide (EPS) is formed, which makes up much of the biofilm matrix. This EPS is also sialylated in the presence of sialic acid, which is present in host tissues. 135 Respiratory infection can be a poly-microbial disease and P. multocida and/or M. haemolytica have readily been isolated from calves infected with H. somni. There is evidence that P. multocida can co-exist within an H. somni biofilm.83 While in a biofilm bacteria are more resistant to host defences and antibiotics.138, 175 Therefore, research on the control of chronic infections should focus on practices to remove the biofilm. Additional details regarding the virulence factors of H. somni have been extensively reviewed.84
Intranasal inoculation or aerosol exposure may result in transient fever and bacteraemia in the absence of clinical signs. Histophilus somni possesses specific adhesins for respiratory tract epithelium, including IbpA, type IV pili (identified from the genome sequence), and likely others, such as LOS. Adherence is the first step in biofilm formation, and mutations in the N-terminus region of ibpA (responsible for adherence) render the bacteria deficient in biofilm formation.136 Adherence to the vascular epithelium may be an initial event in the pathogenesis of disease.75, 121 In fact, the hallmark of H. somni disease is vasculitis.14, 104 Inflammation due to endotoxin and the IbpA Fic motif may enable invasion of the surrounding tissues resulting in bacteraemia and subsequent tissue invasion.25, 59 The Fic motif at the C terminus of IbpA has been shown to cause retraction of respiratory epithelial cells and transmigration of H. somni across the barrier of alveolar cells.183 The nature and severity of clinical signs will depend on the site(s) and extent of such localization. A possible alternative route for organisms invading the central nervous system from the reproductive tract is via Batson’s veins (plexi venosi vertebralis).114, 115 Direct retrograde spread of infection from the orbital or nasal cavities and sinuses to intracranial veins should also possibly be considered as a pathogenic mechanism in TME.
Although syndromes resembling those of the natural disease may be produced by inoculation of the bacterium by several different routes, the results obtained are not always predictable.104 Thrombotic meningoencephalitis may be experimentally reproduced following the intravenous, intracarotid and intracisternal administration of the organisms.75, 104, 126 It has been shown that, while there does seem to be a predilection for the central nervous system following intravenous administration, this is not an inevitable site of localization. Other preferential sites of localization are the cerebrospinal fluid, kidneys and urine with smaller numbers of bacteria being present in other organs.75, 149 Thrombotic meningoencephalitis has not been reproduced by the administration of the bacterium by other routes, such as the intratracheal, intraperitoneal or intramuscular, which supports the hypothesis that bacteraemia is an essential feature in the pathogenesis of the nervous syndrome and probably also of many of the other syndromes.
The pneumonic syndrome has been more successfully reproduced following intratracheal or intrabronchial inoculation of virulent strains of H. somni than it has following nasal inoculation, aerosol exposure or intravenous inoculation.29, 55, 157 As is the case with M. haemolytica, most experimental studies show that aerosol challenge with H. somni alone is not capable of inducing pneumonia, but that other stress factors and/or concomitant infection with viruses or Mycoplasma spp. are required.157 For example, inoculation of even large numbers of pathogenic H. somni into the upper respiratory tract of healthy calves does not result in lower respiratory tract disease, unless the animals are first compromised by challenge with bovine herpes virus I or dexamethasone.40
Abortion has resulted after the inoculation of H. somni into the middle uterine artery and into the amnion114, 115 of pregnant cows as well as by the jugular vein route.176 Severe metritis and abortion may result after intracervical inoculation.75 Introduction of H. somni into the uterus or cervix of non-pregnant cows may cause transient endometritis, cervicitis, vaginitis or vulvitis with persistence of the organism, particularly in the major vestibular glands.97 The organism can adhere to the zona pellucida of the bovine embryo, resulting in embryonal degeneration.165
The primary lesion in the pathological process is considered to be damage to the endothelial cells of small vessels, which predisposes to vasculitis, thrombosis, infarction, ischaemic necrosis and haemorrhage.12, 104, 162 An important feature of infection due to H. somni is the capability of the organism to cause thrombus formation, which may result in septicaemia, disseminated intravascular coagulation (DIC), and TME. As described above, activation of bovine platelets by ChoP expressed on the LOS may trigger the coagulation cascade.14 Initial endothelial damage occurs after many bacteria adhere to the endothelium, a process that is enhanced by the presence of tumour necrosis factor-α.99, 162 Lipooligosaccharide and possibly other components of culture filtrates (possibly peptidoglycan) have been shown to induce endothelial cell apoptosis, resulting in widespread contraction and desquamation of these cells.152 Large areas of subendothelial collagen are thus exposed, presumably initiating the clotting cascade. As a result of retraction of endothelial cells by the Fic motif of IbpA and production of histamine by H. somni, the organisms are allowed direct access to the underlying tissue.104, 132, 162, 183 Bacteria are often observed in the resultant thrombi, and there is an intense neutrophilic response around the affected blood vessels and in the necrotic tissue of the infarcts.162 DIC has been demonstrated in natural cases of H. somni infection and it has been suggested that DIC may play a role in the pathogenesis of the disease.117 Histophilus somni may impair bovine alveolar macrophage and neutrophil phagocytic function,49, 72 possibly due to the network of IbpA on the bacterial surface.27 It has also been established that when bovine neutrophils and mononuclear phagocytes are able to phagocytose opsonised H. somni, they may fail to kill them.32, 50, 51, 180 Because H. somni can multiply and persist in phagocytic cells,103 these cell types may play a role in the pathogenesis of the disease. The bacteria can persist in monocytes in membrane-bound vacuoles, and cytokine activation of these cells fails to eliminate them.51 The mechanism of intracellular survival appears to be due to inhibition of the oxidative burst by phagocytic cells69 and by inhibition of phagosome-lysosome fusion, and not cytoxicity.125 Evasion of killing by neutrophils may also be in part due to the cells undergoing apoptosis.180
Histophilus somni and its outer membrane vesicles, but not purified LOS, have been shown to trigger neutrophil extracellular traps (NETs),65 but it is not clear how effective NETs are in control and clearance of H. somni.
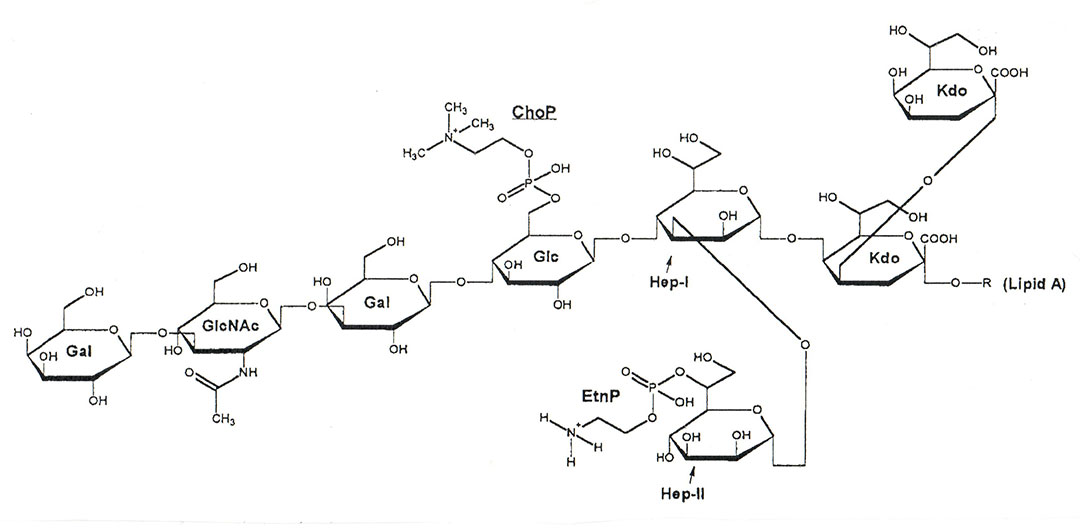
Figure 3 Modified Haworth projection of the predominant structure of H. somni strain 738 LOS. All outer core hexoses are beta-D oriented and both inner core L-glycero-D-manno heptoses are L-alpha-D oriented. Abbreviations: Gal, galactose; GlcNAc, N-acetyl-glucosamine; Glc, glucose; Hep, heptose; Kdo, 3-deoxy-Dmanno-octulosonic acid.

Clinical signs
The most common clinical syndromes currently associated with H. somni involve the respiratory system, joints, and myocardium.57, 123 Other sites affected by H. somni include the central nervous and reproductive systems.75, 97 The organism may also occasionally cause mastitis,62, 66 conjunctivitis,100, 120 otitis,120 and abscessation.146
In North America, the disease is primarily associated with feedlots in older calves and yearlings.41 Clinical signs indicating localization of H. somni in tissues or organs other than the central nervous system may be present59, 75 both in the acute form and in the more protracted or subacute form of the disease. These are represented, for example, by the presence of one or more swollen joints due to arthritis, resulting in lameness and stiffness. When the myocardium is affected, sudden death may result from acute heart failure.59
Thrombotic meningoencephalitis
The TME syndrome is considered to be the outcome of the peracute or acute septicaemic form of the disease. However, in some animals evidence of respiratory disease may precede or accompany the nervous signs.75, 105, 126, 139
The occurrence of sudden death or of animals being found moribund may herald an outbreak of TME.12, 53, 75 In the very early stage animals show a high fever (up to 42 °C) and often profound depression,59 the latter being the most important clinical finding. Other early signs are reluctance to move, stiffness of the legs and knuckling of the fetlocks.59, 75, 92 Evidence of involvement of the nervous system soon widens to include such signs as muscle tremors, blindness, excitement, irritability, circling, ataxia, paresis, paralysis and paddling movements.75 Affected animals generally become comatose in the terminal stages.75, 92, 105
During the course of the disease, the preservation of pupillary light reflexes of the eyes may be absent,171 and ophthalmoscopic examination may reveal the presence of scattered focal retinal haemorrhages with ill-defined borders which, when present, provide a reliable presumptive diagnosis. Hyphaema and hypopyon may also be present.37, 75
Animals suffering from the advanced neurological syndrome invariably die,89 but those treated in the very early stage of clinical disease may recover, although in some that do recover, mild neurological signs may persist.
Respiratory disease syndrome
Apart from the respiratory signs present in some cases of TME, BRD characterized by acute fibrinous pneumonia with or without accompanying pleuritis may be associated with or indicative of H. somni infection. Common clinical signs include depression (indicated by general lack of interest in the environment and drooping of the head and ears), complete or partial inappetence, respiratory signs including tachypnoea and or dyspnoea, mucopurulent nasal discharge, cough, and, in severe cases, extended neck or even open-mouth breathing. Bovine respiratory disease due to H. somni is most common in calves, but may also affect older cattle.18, 130, 140 It is most commonly encountered in calves recently undergoing severe stressors, including weaning, shipping, commingling, and introduction to novel environments and feedstuffs, but can occur later in the feeding period as well. This pattern can be delayed or altered by practice of metaphylaxis (see Mannheimiosis and pneumonic pasteurellosis). It is not possible to distinguish various bacterial agent infections eg. (M. haemolytica, P. multocida, or H. somni) based upon signalment, clinical signs, or history.
Viral infection due to bovine respiratory syncytial virus (BRSV), parainfluenza 3 virus, adenovirus, bovine viral diarrhoea virus, and infectious bovine rhinotracheitis virus may predispose the animals to bacterial infection due to compromising innate host defence mechanisms. In particular, infection of calves with BRSV six days before challenge with H. somni results in more severe disease than either agent alone, possibly due to enhanced IgE production and/or enhanced breakdown of collagen in the lung extracellular matrix. 2, 45 Other bacteria, particularly Mycoplasma spp., M. haemolytica and P. multocida, may increase the severity of the pneumonia due to concurrent or secondary infection.40, 47, 141 In addition to the pneumonic syndrome, H. somni can cause laryngitis59 as well as tracheitis, which results in chronic coughing.75
Histophilus somni has also been associated with pneumonia in water buffalo (Bubalus bubalis) calves in Egypt.86 and American Bison (Bison bison).38
Myocarditis
The significance of myocarditis in histophilosis was not fully recognized until the late 1980s 57, and is now regarded as the most common form of histophilosis resulting in sudden death.122, 123 However, chronic illness may also occur, presenting with depression, lethargy, and poor weight gain. 123 In chronic cases of myocarditis, H. somni is most likely present as a biofilm.136
Reproductive disease syndromes
Histophilus somni may be present in the genital tracts of male and female ruminants without presentation of any clinical signs.123 However, H. somni has been isolated from the genital tract of cows with metritis, endometritis, cervicitis, vaginitis and infertility150, 171 as well as from aborted foetuses and their placentas.140, 151, 171 Spontaneous abortion due to H. somni is generally sporadic rather than occurring as an abortion storm, and usually occurs between the seventh and ninth months of gestation. It is associated with retention of the placenta and severe placentitis.97 The abortion rate may vary from 3 to 12 per cent.171 In one large study from the American Midwest, H. somni was considered the 11th most common cause of bacterial abortion, or 0.23 per cent of all abortions recorded.91, 123
Although H. somni can frequently be isolated from the semen, prepuce, and elsewhere in the urogenital system of bulls without any indication of disease,74 the organism has been associated with a purulent ejaculate.31 Suppurative orchitis associated with H. somni has been reported in calves.94, 113 Histophilus somni has also been cultured from vesicular gland fluid and semen of bulls with vesiculitis.56 However, many isolates from the bovine prepuce are not virulent, are serum-sensitive, and lack virulence factors that enable them to resist host defence mechanisms.28, 143, 184 Histophilus somni can also be a cause of reproductive disease in sheep, particularly rams.123
Other disease syndromes
Spontaneously developing cases of acute to peracute mastitis have been reported in cows due to H. somni.11, 52, 66 The response to antibiotic therapy is variable, with some cases becoming chronic. Acute gangrenous mastitis and chronic mastitis have been produced by inoculation of the organism into the udder.63 In the case of the chronic form of the disease, affected cows shed the bacterium in the milk for up to a year.
Outbreaks of otitis externa have been described in feedlot cattle in Canada,111, 120 and H. somni has been isolated in pure culture and in large numbers from two of the affected animals. Morbidity is generally low, but can reach 10 per cent in some feedlots. The otitis is generally unilateral, but may be bilateral. The affected ear is held lower than normal and there is a copious serous discharge to the exterior. There may or may not be concurrent respiratory disease, but in many outbreaks there is not, and TME has not occurred at the same time in affected herds. Sudden death due to a unilateral suppurative otitis media and interna, and meningitis, has been reported in a heifer.101
A case has been described of pyelonephritis in a cow from which H. somni was cultured.101 The organism has also been isolated from mucopurulent exudate from a urachal abscess in a two-month-old heifer with a distended abdomen.146
Pathology
The pathology and other aspects of disease produced by H. somni have been reviewed by Humphrey and Stephens75 and more recently by O’Toole and Sondgeroth.123 Gross lesions are frequently present in tissues and organ systems other than those of the central nervous system in cases of TME. These include, inter alia, fibrinopurulent polyarthritis and synovitis, rhinitis, sinusitis, necrotizing or ulcerative laryngitis, fibrinonecrotic tracheitis; exudative or interstitial pneumonia;4, 18 serofibrinous pleuritis, pericarditis and peritonitis, enteritis, infarcts in the myocardium57, 88 and kidneys, and regional or generalized lymphadenitis as evidenced by enlargement, congestion, oedema and haemorrhage of the affected lymph nodes.126
Haematological examination of both the naturally occurring and experimentally induced TME reveals that there is initial neutropenia followed by neutrophilia.75
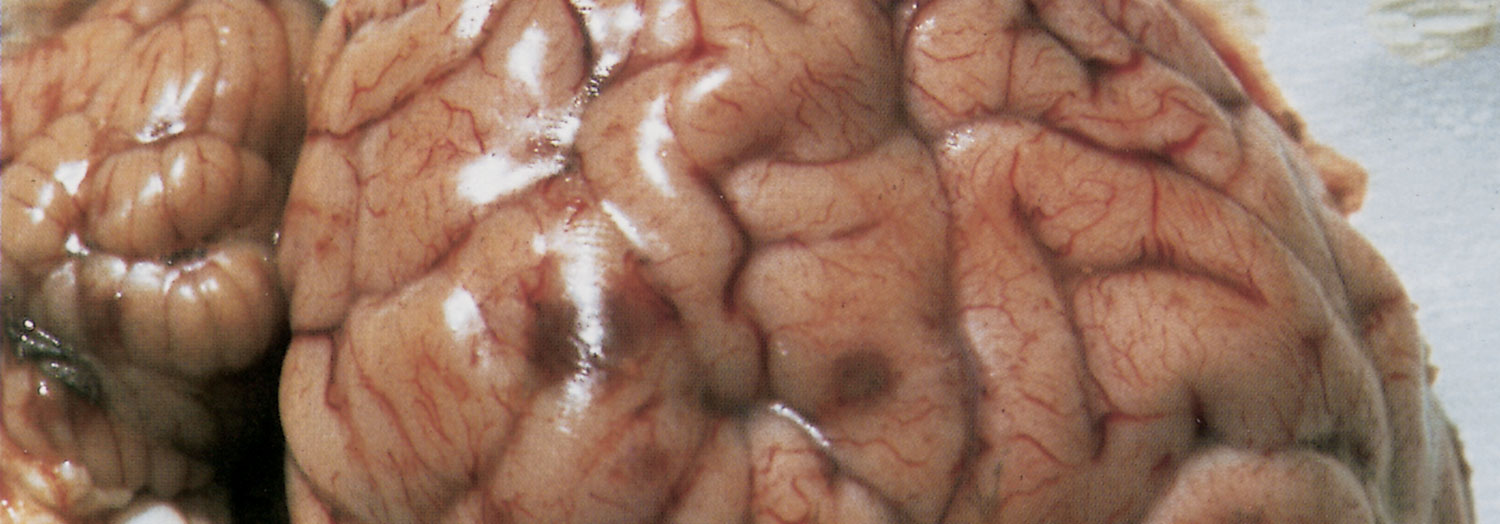
Macroscopically, in animals that have died from TME, the characteristic lesions are multiple infarcts in the central nervous system, which present as reddish-brown foci of haemorrhagic necrosis, varying in diameter from pinpoint to 40 mm (Figure 7).92, 126, 149 The extent and distribution of these infarcts in the brain and spinal cord are variable, but they occur in both the grey and white matter.
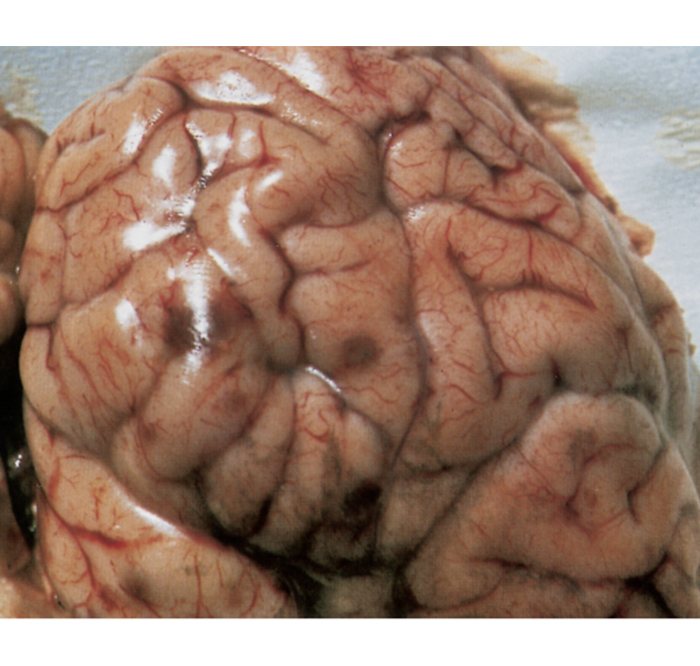
Lesions may be more prevalent in the thalamus, subthalamic nuclei and cerebrum, frequently near the junction of the grey and white matter.126 In some animals, most of the foci are very small (sometimes microscopic) and occur mainly in the spinal cord, while in others only a few very large lesions are present.
Focal fibrinopurulent meningitis may be associated with infarcts, or the meningitis may be more diffuse.126 Fibrinopurulent meningitis in the absence of encephalitic lesions may also occur.85 The cerebrospinal fluid is usually cloudy, yellowish-tinged and contains flocculent debris, and it may coagulate within minutes on exposure to air.126 Analysis of the cerebrospinal fluid may disclose pleocytosis, with neutrophils predominating, increased globulin content, a total protein content greater than 200 mg/100 ml, and lowered glucose content.105
Microscopically, the lesions in TME are typified by severe vasculitis, thrombosis with or without infarction, and a cellular exudate composed almost entirely of neutrophils.126 Such lesions are seen consistently in the brain and cerebral meninges, but may be present in many other tissues, such as the myocardium, kidneys, skeletal musculature, liver, thymus, urinary bladder, and optic tract and retina.75, 88, 105, 126 Large numbers of Gram-negative coccobacilli are often present in some of the smaller thrombosed blood vessels.75, 92, 105 The capability of the H. somni LOS to cause apoptosis of endothelial cells,152 and the IbpA Fic motif to retract epithelial cells, with increased secretion of metalloproteases that damage basement membrane collagen, contribute to vasculitis and dissemination of H. somni.25, 183
Several forms of pneumonia in calves that have suffered from BRD caused by H. somni have been described that are not associated with TME.18, 75 Pneumonia frequently presents as severe, acute fibrino-haemorrhagic bronchopneumonia and fibrinous pleuritis resembling those seen in M. haemolytica infections. Histologically in acute cases there is extensive alveolar and interstitial fibrin deposition, haemorrhage, vascular thrombosis, pulmonary necrosis, and intense alveolar and interstitial leukocyte infiltration. Bryson et al.18 reported that lesions of exudative bronchopneumonia due to H. somni could be divided into two main groups on the basis of histopathological and immunocytochemical features. In one group, cases are dominated by necrotizing bronchiolitis, degeneration and necrosis of airway and alveolar exudates, severe alveolitis with accumulations of degenerate basophilic cells, interstitial inflammatory changes and the widespread distribution of H. somni antigen in airways and alveoli. In the second group, necrotizing and degenerative changes are much less extensive and less severe, and the overall appearance is that of suppurative bronchopneumonia, with H. somni antigen much less widespread within the lungs. No major differences are present between the histopathology of lungs where H. somni was the sole isolate and that of lungs where H. somni was isolated along with other bacterial pathogens. Alveolar oedema, hyaline membrane formation and alveolar epithelial hyperplasia may also be present in caudodorsal lung areas.18 Lung lesions resembling those of the natural disease have been reproduced experimentally by introducing H. somni into the trachea or bronchi of calves.22, 40, 47, 87, 128
Myocarditis is commonly found in association with BRD, and is a common cause of sudden death in feedlot cattle.123 Myocardidis due to H. somni usually affects one or both papillary muscles of the left ventricle.122 During necropsies of cases of BRD or sudden death in cattle, the heart should always be examined for evidence of myocarditis. Myocarditis is a common feature of histophilosis if a thorough necropsy is conducted.109 The bacteria are predominantly present as a biofilm in the myocardium,122 which has been supported by experimental infection.136 The natural state of H. somni is a biofilm,137 which can also occur in the leptomeninges109 and in the respiratory tract.136
There are few studies of the lesions produced by H. somni in the reproductive tract of cattle and in aborted foetuses and their membranes. 75, 97, 114, 115 Gross lesions that have been observed are oedema and necrosis of cotyledons in the placenta, and pronounced diffuse oedema of the foetal extremities. Microscopically, fibrinoid necrosis of arteries of the placenta may be present and the foetus may show vascular necrosis, thrombosis and leukocytosis of the vessels in the brain, lungs, myocardium and renal glomeruli. Infiltration of neutrophils into the alveoli of the foetal lungs and bronchioli may occur, resulting in interstitial pneumonia and myocarditis. The vaginitis, cervicitis and endometritis present in both naturally occurring and experimentally-induced disease seem to be mainly of a purulent nature.30
Diagnosis and differential diagnosis
Since infection by H. somni can present as a variety of nonspecific syndromes, clinically distinguishing these syndromes from others of different aetiology may be difficult.59 Thrombotic meningoencephalitis may be confused clinically with conditions such as viral encephalitides (e.g. rabies, meningoencephalitis caused by bovine herpesvirus-1), heartwater, cerebral redwater, cerebral theileriosis, listeriosis, sporadic bovine encephalomyelitis caused by Chlamydophila pecorum, bacterial meningitis caused, for example, by M. haemolytica or P. multocida, pituitary or cerebral abscessation, polioencephalomalacia (or cerebrocortical necrosis) caused by a thiamine deficiency, sulphur intoxication, and other intoxications, such as lead poisoning, botulism, diplodiosis, Aspergillus clavatus mycotoxicosis, and pesticide poisoning.
Two important clinical findings that may help to differentiate histophilosis from other diseases are the very profound depression or somnolence59 (hence the term ‘sleeper syndrome’) that occurs in the early stages of TME, and the focal retinal haemorrhages visible on ophthalmoscopic examination which, when present, provide a reliable presumptive diagnosis.37, 75
Due to the potential for multi-systemic infection by H. somni, necropsies should be thorough. Confirmation of TME depends on histopathological examination and bacteriological culture of the affected tissues. However, bacteriological culture of the blood and cerebrospinal fluid alone in clinical cases yields H. somni inconsistently.59, 149 The organism does not survive for a prolonged period in commonly used transport media.17, 59 Swabs for bacterial culture should be kept moist and, if possible, refrigerated, and rapidly transported to the laboratory59 for culture within 24 hours. Nonculture methods such as PCR or immunohistochemistry are useful adjuncts to culture. Serology may be useful, but must be interpreted in light of vaccination history.
From a pathological viewpoint the haemorrhagic infarcts in the central nervous system in TME are considered to be characteristic and are macroscopically visible in the majority of cases. Thrombosis and infarcts are also present in the central nervous system of animals suffering from cerebral theileriosis. These two diseases can be differentiated by the microscopic detection of Koch’s bodies in the case of cerebral theileriosis in smears made from the contents of thrombosed blood vessels or by histopathology (see Turning sickness).
The ‘sudden death’ syndrome in histophilosis, resulting in death from acute heart failure following focal myocardial infarction, should be differentiated from other possible causes of this syndrome in feedlot cattle, such as acute pneumonic mannheimiosis/pasteurellosis, blackleg (clostridial disease), ruminal acidosis and tympany, polioencephalomalacia, and poisonings (e.g. urea and ionophore poisonings).
The respiratory disease syndrome caused by H. somni should primarily be differentiated from pneumonia caused by M. haemolytica, P. multocida, Mycoplasma spp. and other organisms involved in the BRD complex. However, this cannot readily be achieved on clinical or pathological grounds. Therefore, a definitive diagnosis rests on specific detection of the bacteria, bacterial antigens, or their DNA from pneumonic lesions.
The infertility syndrome in cows caused by H. somni should be differentiated from the many other potential causes, including bacterial (Campylobacter, Arcanobacterium), protozoal (Tritrichomonas), fungal and viral (bovine herpesvirus 1, bovine viral diarrhoea virus) agents. Bacteriological culture of H. somni from the bovine genital tract may be difficult due to faecal contamination and culture overgrowth by Escherichia coli and Proteus spp. Therefore, a protected swabbing technique is advocated.97
Due to the slow growth of H. somni in vitro it may be outgrown by other bacteria, e.g. M. haemolytica or P. multocida in the case of samples from pneumonic lesions. Use of a selective medium is therefore advocated, such as one that incorporates vancomycin (5 µg/ml), neomycin (5 µg/ml), nystatin (100 IU/ml), cyclohexamide (100 µg/ml) and thiamine monophosphate (1 µg/ml) into 5 per cent horse blood agar.145 Nonetheless, bacterial culture is a relatively insensitive diagnostic method compared with more recent molecular methods. A variety of immunological and DNA tests have been developed for the detection of H. somni. These include techniques using monoclonal164 or polyclonal158 antibodies, immunoperoxidase staining,58 in situ hybridization,136 various PCR formats,6, 107, 160 and an optical fibre biosensor for DNA detection.13 A comparison of several of these methods, including bacterial culture, showed that PCR or a biosensor for DNA were the most sensitive methods, whether performed on swabs from the cut surface of the lung, on bronchial content or on growth from agar plates.13, 156
The use of classical serologic tests (detection of antibodies) for the diagnosis of clinical cases of histophilosis has been equivocal. In herds in which TME has been diagnosed, more than 50 per cent of normal cattle may be seropositive, indicating that while TME is generally sporadic, most animals that become infected do not show evidence of the disease. Even rising antibody titres in successive examinations do not necessarily provide meaningful evidence, as this phenomenon has also been observed in asymptomatic cattle.59, 75 In addition, when interpreting serological results, the prior vaccination history of the animals should be taken into consideration.
Another problem in the interpretation of serological data is lack of specificity, as antibodies to H. somni whole cells may cross-react with other bacteria, such as other Haemophilus spp., Actinobacillus lignieresii, Listeria monocytogenes, Bordetella bronchiseptica, and others.75, 149
Serological tests that use whole cells as antigen include the microagglutination test, tube agglutination tests, complement fixation test, and enzyme-linked immunosorbent assay (ELISA).59, 75 However, ELISA assays that utilize IbpA106, 181 or the biofilm EPS124 as the diagnostic antigen are much more specific for detecting diseased animals, particularly in chronic infections, as specific antibodies may require at least 2 weeks to be detected.
Control
In vitro tests indicate that H. somni is generally susceptible to most commonly used antimicrobials.75, 174 Histophilus somni has been shown to be sensitive in vitro to ceftiofur,133 tilmicosin,118 florfenicol,35 danofloxacin,46 enrofloxacin, ciprofloxacin, and marbofloxacin.95, 129 However, in a large study of 745 BRD pathogens isolated in Alberta, Canada, 66.7 per cent of H. somni isolates were resistant to at least one antibiotic, whereas 100 per cent of all other BRD bacterial pathogens were resistant to at least one antimicrobial. Multi-drug resistance was present in 6.7 per cent of H. somni isolates.7 Sporadic occurrences of resistance to ampicillin, tetracycline, tilmicosin, erythromycin, streptomycin, neomycin, kanamycin, framycetin, lincomycin, spectinomycin and sulphonamides have been reported.52, 86, 172, 174 Such resistance may, in part, be due to “silent genes” that have the potential to be transcribed and enhance resistance, but most often are not transcribed. For example, the genome of H. somni contains genomic islands that further contain homologues of tetA and romA (a multi-drug resistance gene) genes responsible for some tetracycline resistance, although most isolates are susceptible to tetracycline.116 Furthermore, the tetH gene was found in all H. somni isolates examined from an Alberta feedlot.34
Studies from Denmark and across Europe showed that H. somni and other respiratory disease pathogens were overall highly susceptible to most antibiotics, with the exception of tetracycline.1, 39 Histophilus somni isolates from Australia were found to be highly clonal and also highly susceptible to most antibiotics.48 A study from the Iowa State University Veterinary Diagnostic Laboratory that examined 1251 BRD isolates showed that more isolates, including H. somni, were more resistant to antimicrobial agents if the cattle had previously been treated with antibiotics, and the percentage of resistant isolates increased as the number of treatments with antimicrobial agents increased.108
In North America, where the disease is most prevalent, it is a standard recommendation in the event of an outbreak to increase surveillance and to institute immediate treatment with an antimicrobial. Mass medication with antibiotics in the feed, or the treatment of every animal in the group that contains affected animals with a long-acting antibiotic, has yielded variable results.59 In animals suffering from TME, treatment must commence as early in the course of the disease as possible, as once the neurological signs are well established, treatment is generally not successful. Oxytetracycline injected intravenously early in the course of TME is successful in preventing mortality.59, 92
One large trial showed that additional mass prophylactic medication of calves with long-acting oxytetracycline on days 8, 11 or 17 after arrival in the feedlot did not reduce the risk of histophilosis mortality.169
Several H. somni bacterins, alone or in combination with M. haemolytica and respiratory viral pathogens, are commercially available in North America, and their use has yielded beneficial, although inconsistent, results.54, 119, 131, 167 Bacterins may be responsible for the reduced incidence of TME, but appear to have little efficacy against BRD. Vaccination of beef calves at three and five weeks of age with a combination M. haemolytica-H. somni bacterin vaccine failed to reduce the prevalence of enzootic pneumonia.170 The lack of consistent evidence for the effectiveness of current vaccines has been supported by a review of the vaccine literature.102 However, a strong immune response with high serum antibody levels is not necessarily indicative of protection.147 The lack of consistent efficacy may be related to phase variation in the LOS,79 to biofilm formation,136 and intracellular persistence (see below). Commercial vaccines containing H. somni and bovine respiratory syncytial virus antigens have been reported to have a high rate of adverse reactions.131 Such reactions may be due to Type 1 (immediate) hypersensitivity, as commercial H. somni bacterins have been shown to elicit an IgE response.131 Some H. somni vaccine research has concentrated mainly on defining unique surface antigens. A leading candidate at this time is the IbpA region containing the FIC motif, described above.43, 44
Immunoglobulin G2 has been found in the lumen of the bovine uterus after intramuscular immunization and may be important in local immunological protection in the bovine reproductive tract.19 However, vaccination with H. somni bacterins has shown no efficacy against this organism in protection against reproductive system disease.67 Vaccination of cows prepartum with a combined M. haemolytica and H. somni bacterin vaccine results in passive antibody titres in calves up to two months of age, but it is not known whether there is any protective effect.167
A major limitation of bacterins and subunit vaccines is that they induce primarily a Th2 or antibody-mediated immune response. However, H. somni has long been known to be capable of surviving within phagocytic cells.32, 103, 125 As a result, H. somni may be considered a persistent intracellular pathogen. Intracellular bacteria are not susceptible to antibody-mediated immunity, which may be why bacterins have had some success against TME, but little to none against pneumonia and myocarditis. Therefore, a Th1 cellular immune response, which can be stimulated by a live, attenuated strain, may be more successful, but has not yet been tested for the various syndromes of histophilosis. When in a biofilm, the bacteria are also more resistant to host defence mechanisms. Therefore, when one considers the host resistance mechanisms of H. somni (LOS phase variation, biofilm formation, IbpA FIC toxicity, and intracellular survival) the optimal vaccine would have to be designed to overcome each of these resistance mechanisms.
References
- AARESTRUP, F. M., SEYFARTH, A. M. & ANGEN, Ø., 2004. Antimicrobial susceptibility of Haemophilus parasuis and Histophilus somni from pigs and cattle in Denmark. Veterinary Microbiology, 101, 143-146.
- AGNES, J. T., ZEKARIAS, B., SHAO, M., ANDERSON, M. L., GERSHWIN, L. J. & CORBEIL, L. B., 2013. Bovine respiratory syncytial virus and Histophilus somniinteraction at the alveolar barrier. Infection and Immunity, 81, 2592-2597.
- AKHTAR, S., FARVER, T. B. & RIEMANN, H. P., 1997. A sero-epidemiological study of Haemophilus somni infection in dairy cattle. Veterinary Research Communications, 21, 229-239.
- ANDREWS, J. J., ANDERSON, T. D., SLIFE, L. N. & STEVENSON, G. W., 1985. Microscopic lesions associated with the isolation of Haemophilus somni from pneumonic bovine lungs. Veterinary Pathology, 22, 131-136.
- ANGEN, Ø., 2016. Taxonomy of Histophilus somni. Current Topics in Microbiology and Immunology, 396, 1-14.
- ANGEN, Ø., AHRENS, P. & TEGTMEIER, C., 1998. Development of a PCR test for identification of Haemophilus somni in pure and mixed cultures. Veterinary Microbiology, 63, 39-48.
- ANHOLT, R. M., KLIMA, C., ALLAN, N., MATHESON-BIRD, H., SCHATZ, C., AJITKUMAR, P., OTTO, S. J., PETERS, D., SCHMID, K., OLSON, M., MCALLISTER, T. & RALSTON, B., 2017. Antimicrobial susceptibility of bacteria that cause bovine respiratory disease complex in Alberta, Canada. Frontiers in Veterinary Science, 4, 207.
- ANONYMOUS., 2019. Histophilus somni infection in cattle. Veterinary Record, 185, 45.
- APPUHAMY, S., LOW, J. C., COOTE, J. G. & PARTON, R., 1998. PCR methods and plasmid profile analysis for characterisation of Histophilus ovis strains. Journal of Medical Microbiology, 47, 987-992.
- APPUHAMY, S., PARTON, R., COOTE, J. G. & GIBBS, H. A., 1997. Genomic fingerprinting of Haemophilus somni by a combination of PCR methods. Journal of Clinical Microbiology, 35, 288-291.
- ARMSTRONG, K. R., OSBORNE, A. D. & JANZEN, E. D., 1986. Haemophilus somni mastitis in a dairy cow. Canadian Veterinary Journal, 27, 211-212.
- BAILIE, W. E., ANTHONY, H. D. & WEIDE, K. D., 1966. Infectious thromboembolic meningoencephalitis (sleeper syndrome) in feedlot cattle. Journal of the American Veterinary Medical Association, 148, 162-166
- BANDARA, A. B., ZUO, Z., MCCUTCHEON, K., RAMACHANDRAN, S., HEFLIN, J. R. & INZANA, T. J., 2018. Identification of Histophilus somni by a nanomaterial optical fiber biosensor assay. Journal of Veterinary Diagnostic Investigation, 30, 821-829.
- BEHLING-KELLY, E., RIVERA-RIVAS, J. & CZUPRYNSKI, C. J., 2016. Interactions of Histophilus somniwith Host Cells. Current Topics in Microbiology and Immunology, 396, 71-87.
- BEKER, M., ROSE, S., LYKKEBO, C. A. & DOUTHWAITE, S., 2018. Integrative and conjugative elements (ICEs) in Pasteurellaceae species and their detection by multiplex PCR. Frontiers in Microbiology, 9, 1329.
- BOOKER, C. W., GUICHON, P. T., JIM, G. K., SCHUNICHT, O. C., HARLAND, R. J. & MORLEY, P. S., 1999. Seroepidemiology of undifferentiated fever in feedlot calves in western Canada. Canadian Veterinary Journal, 40, 40-48.
- BREWER, R. A., CORBEL, M. J. & STUART, F. A., 1985. Development of improved methods for the transport and isolation of Haemophilus somni. Research in Veterinary Science, 39, 299-306.
- BRYSON, D. G., BALL, H. J., MCALISKEY, M., MCCONNELL, W. & MCCULLOUGH, S. J., 1990. Pathological, immunocytochemical and microbiological findings in calf pneumonias associated with Haemophilus somni infection. Journal of Comparative Pathology, 103, 433-445.
- BUTT, B. M., BESSER, T. E., SENGER, P. L. & WIDDERS, P. R., 1993. Specific antibody to Haemophilus somni in the bovine uterus following intramuscular immunization. Infection and Immunity, 61, 2558-2562.
- CHALLACOMBE, J. F., DUNCAN, A. J., BRETTIN, T. S., BRUCE, D., CHERTKOV, O., DETTER, J. C., HAN, C. S., MISRA, M., RICHARDSON, P., TAPIA, R., THAYER, N., XIE, G. & INZANA, T. J., 2007. Complete genome sequence of Haemophilus somnus (Histophilus somni) strain 129Pt and comparison to Haemophilus ducreyi 35000HP and Haemophilus influenzae Rd. Journal of Bacteriology, 189, 1890-1898.
- CIRONE, F., PADALINO, B., TULLIO, D., CAPOZZA, P., LO SURDO, M., LANAVE, G. & PRATELLI, A., 2019. Prevalence of pathogens related to bovine respiratory disease before and after transportation in beef steers: Preliminary Results. Animals (Basel), 9(12), e1093.
- CONFER, A. W., SNIDER, T. A., TAYLOR, J. D., MONTELONGO, M. & SORENSEN, N. J., 2016. Clinical disease and lung lesions in calves experimentally inoculated with Histophilus somni five days after metaphylactic administration of tildipirosin or tulathromycin. American Journal of Veterinary Research, 77, 358-66.
- CORBEIL, L. B., 1990. Molecular aspects of some virulence factors of Haemophilus somni. Canadian Journal of Veterinary Research, 54, S57–S62.
- CORBEIL, L. B., 2007. Histophilus somnihost-parasite relationships. Animal Health Research Reviews, 8, 151-160.
- CORBEIL, L. B., 2016a. Histophilus somni: Biology, Molecular Basis of Pathogenesis, and Host Immunity. Current Topics in Microbiology and Immunology, Springer, Switzerland, 396.
- CORBEIL, L. B., 2016b. Histophilus somni: Biology, Molecular Basis of Pathogenesis, and Host Immunity. Current Topics in Microbiology and Immunology, Springer, Switzerland, 396.
- CORBEIL, L. B., BASTIDA-CORCUERA, F. D. & BEVERIDGE, T. J., 1997. Haemophilus somni immunoglobulin binding proteins and surface fibrils. Infection and Immunity, 65, 4250-4257.
- CORBEIL, L. B., BLAU, K., PRIEUR, D. J. & WARD, A. C., 1985. Serum susceptibility of Haemophilus somnusfrom bovine clinical cases and carriers. Journal of Clinical Microbiology, 22, 192-198.
- CORBEIL, L. B., WIDDERS, P. R., GOGOLEWSKI, R., ARTHUR, J., INZANA, T. J. & WARD, A. C., 1986. Haemophilus somnus: Bovine Reproductive and Respiratory Disease. Canadian Veterinary Journal, 27, 90-93.
- CORBEIL, L. B., WIDDERS, P. R., GOGOLEWSKI, R., INZANA, I. & WARD, A. C. S., 1986. Haemophilus somni: Bovine reproductive and respiratory disease. Canadian Veterinary Journal, 27, 90-93.
- CORBOZ, L. & WILD, P., 1981. Epidemiologie der Haemophilus somni-Infektion beim Rind: Vergleich von Stämmen in der Polyacrylamidgel-Elektrophorese (PAGE). Schweizer Archiv für Tierheilkunde, 123, 79-88.
- CZUPRYNSKI, C. J. & HAMILTON, H. L., 1985. Bovine neutrophils ingest but do not kill Haemophilus somni in vitro. Infection and Immunity, 50, 431-436.
- CZUPRYNSKI, C. J., LEITE, F., SYLTE, M., KUCKLEBURG, C., SCHULTZ, R., INZANA, T., BEHLING-KELLY, E. & CORBEIL, L. B., 2004. Complexities of the pathogenesis of Mannheimia haemolytica and Haemophilus somnus infections: challenges and potential opportunities for prevention? Animal Health Research Reviews, 5, 277-282.
- D'AMOURS, G. H., WARD, T. I., MULVEY, M. R., READ, R. R. & MORCK, D. W., 2011. Genetic diversity and tetracycline resistance genes of Histophilus somni. Veterinary Microbiology, 150, 362-372.
- DE CRAENE, B. A., DEPREZ, P., D’HAESE, E., NELIS, H. J., VAN DEN BOSSCHE, W. & DE LEENHEER, A. P., 1997. Pharmacokinetics of florfenicol in cerebrospinal fluid and plasma of calves. Antimicrobial Agents and Chemotherapy, 41, 1991–1995
- DEWEY, K. J. & LITTLE, P. B., 1984. Environmental survival of H. somni and influence of secretions and excretions. Canadian Journal of Comparative Medicine, 48, 23-26.
- DUKES, T. W., 1971. The ocular lesions in thromboembolic meningoencephalitis (TEME) of cattle. Canadian Veterinary Journal, 12, 180-182.
- DYER, N. W., 2001. Haemophilus somnus bronchopneumonia in American Bison (Bison bison). Journal of Veterinary Diagnostic Investigation, 13, 419-421.
- EL GARCH, F., DE JONG, A., SIMJEE, S., MOYAERT, H., KLEIN, U., LUDWIG, C., MARION, H., HAAG-DIERGARTEN, S., RICHARD-MAZET, A., THOMAS, V. & SIEGWART, E., 2016. Monitoring of antimicrobial susceptibility of respiratory tract pathogens isolated from diseased cattle and pigs across Europe, 2009-2012: VetPath results. Veterinary Microbiology, 194, 11-22.
- ELSWAIFI, S. F., SCARRATT, W. K. & INZANA, T. J., 2012. The role of lipooligosaccharide phosphorylcholine in colonization and pathogenesis of Histophilus somniin cattle. Veterinary Research, 43, 49.
- FECTEAU, G. & GEORGE, L. W., 2004. Bacterial meningitis and encephalitis in ruminants. Veterinary Clinics of North American Food Animal Practice, 20, 363-377.
- FUSSING, V. & WEGENER, H. C., 1993. Characterization of bovine Haemophilus somni by biotyping, plasmid profiling, REA-patterns and ribotyping. Zentralblatt für Bakteriologie, 279, 60-74.
- GEERTSEMA, R. S., WORBY, C., KRUGER, R. P., TAGAWA, Y., RUSSO, R., HERDMAN, D. S., LO, K., KIMBALL, R. A., DIXON, J. & CORBEIL, L. B., 2008. Protection of mice against Histophilus somnisepticemia by vaccination with recombinant immunoglobulin binding protein subunits. Vaccine. 26, 4506-4512.
- GEERTSEMA, R. S., ZEKARIAS, B., LA FRANCO SCHEUCH, L., WORBY, C., RUSSO, R., GERSHWIN, L. J., HERDMAN, D. S., LO, K. & CORBEIL, L. B., 2011. IbpA DR2 subunit immunization protects calves against Histophilus somnipneumonia. Vaccine, 29, 4805-4812.
- GERSHWIN, L. J., BERGHAUS, L. J., ARNOLD, K., ANDERSON, M. L. & CORBEIL, L. B., 2005. Immune mechanisms of pathogenetic synergy in concurrent bovine pulmonary infection with Haemophilus somnus and bovine respiratory syncytial virus. Veterinary Immunology and Immunopathology, 107, 119-130.
- GILES, C. J., MAGONIGLE, R. A., GRIMSHAW, W. T., TANNER, A. C., RISK, J. E., LYNCH, M. J. & RICE, J. R., 1991. Clinical pharmacokinetics of parenterally administered danofloxacin in cattle. Journal of Veterinary Pharmacology and Therapeutics, 14, 400-410.
- GOGOLEWSKI, R. P., LEATHERS, C. W., LIGGITT, H. D. & CORBEIL, L. B., 1987. Experimental Haemophilus somnuspneumonia in calves and immunoperoxidase localization of bacteria. Veterinary Pathology, 24, 250-256.
- GOLDSPINK, L. K., MOLLINGER, J. L., BARNES, T. S., GROVES, M., MAHONY, T. J. & GIBSON, J. S., 2015. Antimicrobial susceptibility of Histophilus somni isolated from clinically affected cattle in Australia. Veterinary Journal, 203, 239-243.
- GOMIS, S. M., GODSON, D. L., BESKORWAYNE, T., WOBESER, G. A. & POTTER, A. A., 1997. Modulation of phagocytic function of bovine mononuclear phagocytes by Haemophilus somni. Microbial Pathogenesis, 22, 13-21.
- GOMIS, S. M., GODSON, D. L., WOBESER, G. A. & POTTER, A. A., 1997. Effect of Haemophilus somni on nitric oxide production and chemiluminescence response of bovine blood monocytes and alveolar macrophages. Microbial Pathogenesis, 23, 327-333.
- GOMIS, S. M., GODSON, D. L., WOBESER, G. A. & POTTER, A. A., 1998. Intracellular survival of Haemophilus somni in bovine blood monocytes and alveolar macrophages. Microbial Pathogenesis, 25, 227-235.
- GREER, D., MCCONNELL, W. & BALL, H., 1989. Isolation of Haemophilus somni from bovine milk. Veterinary Record, 125, 381-382.
- GRINER, L. A., JENSEN, R. & BROWN, W. W., 1956. Infectious embolic meningo-encephalitis in cattle. Journal of the American Veterinary Medical Association, 129, 417-421
- GROOM, S. C. & LITTLE, P. B., 1988. Effects of vaccination of calves against induced Haemophilus somni pneumonia. American Journal of Veterinary Research, 49, 793-800.
- GROOM, S. C., LITTLE, P. B. & ROSENDAL, S., 1988. Virulence differences among three strains of Haemophilus somni following intratracheal inoculation of calves. Canadian Journal of Veterinary Research, 52, 349-354.
- GROTELUESCHEN, D. M., MORTIMER, R. G. & ELLIS, R. P., 1994. Vesicular adenitis syndrome in beef bulls. Journal of the American Veterinary Medical Association, 205, 874-877.
- GUICHON, P. T., PRITCHARD, J. & JIM, G. K., 1988. Haemophilus somni myocarditis in a feedlot steer. Canadian Veterinary Journal, 29, 1012-1013.
- HARITANI, M., NAKAZAWA, M., HASHIMOTO, K., NARITA, M., TAGAWA, Y. & NAKAGAWA, M., 1990. Immunoperoxidase evaluation of the relationship between necrotic lesions and causative bacteria in lungs of calves with naturally acquired pneumonia. American Journal of Veterinary Research, 51, 1975-1979.
- HARRIS, F. W. & JANZEN, E. D., 1989. The Haemophilus somni complex (Haemophilosis): A review. Canadian Veterinary Journal, 30, 816-822
- HASE, M., 1984. Outbreaks of Haemophilus somni infection in cattle in Kagoshima Prefecture. Journal of the Japan Veterinary Medical Association, 37, 431-435.
- HAZIROGLU, R., ERDEGER, J., GULBAHAR, M. Y. & KUL, O., 1997. Association of Pasteurella haemolytica, Pasteurella multocida and Haemophilus somni with pneumonia in calves. Deutsche Tierärztliche Wochenschrift, 104, 150-153.
- HAZLETT, M. J., LITTLE, P. B. & BARNUM, D. A., 1983. Experimental production of mastitis with Haemophilus somni in the lactating bovine mammary gland. Canadian Veterinary Journal, 24, 135.
- HAZLETT, M. J., LITTLE, P. B., BARNUM, D. A., MAXIE, M. G., LESLIE, K. E. & MILLER, R. B., 1985. Haemophilus somni: Investigations of its potential role in bovine mastitis. American Journal of Veterinary Research, 46, 2229-2234.
- HEADLEY, S. A., PEREIRA, A. H. T., BALBO, L. C., DI SANTIA, G. W., BRACARENSE, A. P. F. R. L., FILHO, L. F. C. C., SCHADE, J., OKANO, W., PEREIRA, P. F. V., MOROTTI, F., PRETO-GIORDANO, L. G., MARCASSO, R. A., ALFIERI, A. F., LISBÔA, J. A. N. & ALFIERI, A. A., 2018. Histophilus somni-associated syndromes in sheep from Southern Brazil. Brazilian Journal of Microbiology, 49, 591-600.
- HELLENBRAND, K. M., FORSYTHE, K. M., RIVERA-RIVAS, J. J., CZUPRYNSKI, C. J. & AULIK, N. A., 2013. Histophilus somnicauses extracellular trap formation by bovine neutrophils and macrophages. Microbial Pathogenesis, 54, 67-75.
- HIGGINS, R., MARTIN, J. R., LAROUCHE, Y. & GOYETTE, G., 1987. Mastitis caused by Haemophilus somni in a dairy cow. Canadian Veterinary Journal, 28, 117-118.
- HJERPE, C. A., 1990. Bovine vaccines and herd vaccination programs. Veterinary Clinics of North America: Food Animal Practice, 6, 171-260.
- HOERLEIN, A. B., GOTO, K. & YOUNG, S., 1973. Haemophilus somni agglutinins in cattle. Journal of the American Veterinary Medical Association, 163, 1375-1377.
- HOWARD, M. D., BOONE, J. H., BUECHNER-MAXWELL, V., SCHURIG, G. G. & INZANA, T. J., 2004. Inhibition of bovine macrophage and polymorphonuclear leukocyte superoxide anion production by Haemophilus somnus. Microbial Pathogenesis, 37, 263-271.
- HOWARD, M. D., COX, A. D., WEISER, J. N., SCHURIG, G. G. & INZANA, T. J., 2000. Antigenic diversity in Haemophilus somnus lipooligosaccharide and investigation of the phosphorylcholine epitope. Journal of Clinical Microbiology, 38, 4412-4419.
- HOWARD, M. D., WILLIS, L., WAKARCHUK, W., ST. MICHAEL, F., COX, A., HORNE, W. T., HONTECILAS, R., BASSAGANYA-RIERA, J., LORENZ, E. & INZANA, T. J., 2011. Genetics and molecular specificity of sialylation of Histophilus somni lipooligosaccharide (LOS) and the effect of LOS sialylation on Toll-like receptor-4 signaling. Veterinary Microbiology, 153, 163-172.
- HUBBARD, R. D., KAEBERLE, M. L., ROTH, J. A. & CHIANG, Y. W., 1986. Haemophilus somni-induced interference with bovine neutrophil functions. Veterinary Microbiology, 12, 77-85.
- HUMPHREY, J. D., LITTLE, P. B., BARNUM, D. A., DOIG, P. A., STEPHENS, L. R. & THORSEN, J., 1982. Occurrence of ‘Haemophilus somni’ in bovine semen and in the prepuce of bulls and steers. Canadian Journal of Comparative Medicine, 46, 215-217.
- HUMPHREY, J. D., LITTLE, P. B., STEPHENS, L. R., BARNUM, D. A., DOIG, P. A. & THORSEN, J., 1982. Prevalence and distribution of Haemophilus somni in the bovine reproductive tract. American Journal of Veterinary Research, 43, 791-795.
- HUMPHREY, J. D. & STEPHENS, L. R., 1983. ‘Haemophilus somni’: A review. The Veterinary Bulletin, 53, 987-1004.
- INZANA, T. J., BALYAN, R. & HOWARD, M. D., 2012. Decoration of Lipooligosaccharide with N-acetyl-5-neuraminic acid contributes to Histophilus somni virulence and resistance to host defenses. Veterinary Microbiology, 161, 113-121.
- INZANA, T. J. & CORBEIL, L. B., 1987. Development of a defined medium for Haemophilus somnusisolated from cattle. American Journal of Veterinary Research, 48, 366-369.
- INZANA, T. J., GLINDEMANN, G., COX, A. D., WAKARCHUK, W. & HOWARD, M. D., 2002. Incorporation of N-acetylneuraminic acid into Haemophilus somnus lipooligosaccharide (LOS): Enhancement of resistance to serum and reduction of LOS antibody binding. Infection and Immunity, 70, 4870-4879.
- INZANA, T. J., GOGOLEWSKI, R. & CORBEIL, L. B., 1992. Phenotypic phase variation in Haemophilus somnus lipooligosaccharide during bovine pneumonia and after in vitro passage. Infection and Immunity, 60, 2943-2951.
- INZANA, T. J., GOGOLEWSKI, R. P. & CORBEIL, L. B., 1992. Phenotypic phase variation in Haemophilus somni lipooligosaccharide during bovine pneumonia and after in vitro passage. Infection and Immunity, 60, 2943-2951
- INZANA, T. J., HENSLEY, J., MCQUISTON, J., LESSE, A. J., CAMPAGNARI, A. A., BOYLE, S. M. & APICELLA, M. A., 1997. Phase variation and conservation of lipooligosaccharide epitopes in Haemophilus somnus. Infection and Immunity, 65, 4675-4681.
- INZANA, T. J., IRITANI, B., GOGOLEWSKI, R. P., KANIA, S. A. & CORBEIL, L. B., 1988. Purification and characterization of lipooligosaccharides from four strains of Haemophilus somni. Infection and Immunity, 56, 2830-2837.
- INZANA, T. J., PETRUZZI, B. J. & DICKERMAN, A., 2019. Poly-microbial biofilm interaction by Histophilus somni and Pasteurella multocida in vitro and in vivo, and their effect on host response. FEMS 2019, 8th Congress of European Microbiologists. Glasgow, Scotland. Abstract # PM297.
- INZANA, T. J. & THOMAS, J., 2016. Histophilus somni: Biology, Molecular Basis of Pathogenesis, and Host Immunity. Current Topics in Microbiology and Immunology. Springer, London, 396.
- ISHIKAWA, Y., TSUKUDA, S., NAKAJIMA, Y. & OHSHIMA, K., 1984. A typical nervous lesion in Haemophilus somni infection of cattle. Cornell Veterinarian, 74, 349-353.
- ISMAIL, M., 1991. Haemophilus somni as a bacterial cause of pneumonia in buffalo calves in Egypt. Archiv für Experimentelle Veterinarmedizin, 45, 161-164.
- JACKSON, J. A., ANDREWS, J. J. & HARRIS, J. W., 1987. Experimental Haemophilus somni pneumonia in calves. Veterinary Pathology, 24, 129-134.
- JANZEN, E. D., 1987. Myocardial lesions caused by H. somni in cattle. Canadian Veterinary Journal, 28, 208.
- KENNEDY, P. C., BIBERSTEIN, E. L., HOWARTH, J. A., FRAZIER, L. M. & DUNGWORTH, D. L., 1960. Infectious meningo-encephalitis in cattle, caused by a Haemophilus-like organism. American Journal of Veterinary Research, 21, 403-409.
- KILIAN, M. & BIBERSTEIN, E. L., 1984. Genus Haemophilus. In: KRIEG, N. R. & HOLT, J. G., (eds.). Bergey’s Manual of Systemic Bacteriology. Baltimore, London: Williams & Wilkins.
- KIRKBRIDE, C. A., 1993. Bacterial agents detected in a 10-year study of bovine abortions and stillbirths. Journal of Veterinary Diagnostic Investigation, 5, 64-68.
- KITCHING, J. P., BISHOP, G. C. & MAPHAM, P. H., 1985. Thromboembolic meningoencephalitis diagnosed in Natal. Journal of the South African Veterinary Association, 56, 97-98.
- KLIMA, C. L., HOLMAN, D. B., RALSTON, B. J., STANFORD, K., ZAHEER, R., ALEXANDER, T. W. & MCALLISTER, T. A., 2019. Lower respiratory tract microbiome and resistome of bovine respiratory disease mortalities. Microbial Ecology, 78, 446-456.
- KREPLIN, C., 1991. Suppurative orchitis associated with Haemophilus somni in a calf. Canadian Veterinary Journal, 32, 567.
- KROEMER, S., GALLAND, D., GUÉRIN-FAUBLÉE, V., GIBOIN, H. & WOEHRLÉ-FONTAINE, F., 2012. Survey of marbofloxacin susceptibility of bacteria isolated from cattle with respiratory disease and mastitis in Europe. Veterinary Record, 170, 53.
- KUCKLEBURG, C. J., ELSWAIFI, S. F., INZANA, T. J. & CZUPRYNSKI, C. J., 2007. Expression of phosphorylcholine by Histophilus somni induces bovine platelet aggregation. Infection and Immunity, 75, 1045-1049.
- KWIECIEN, J. M. & LITTLE, P. B., 1991. Histophilus somni and reproductive disease in the cow: A review. Canadian Veterinary Journal,32, 595-601.
- KWIECIEN, J. M. & LITTLE, P. B., 1992. Isolation of pathogenic strains of Haemophilus somni from the female bovine reproductive tract. Canadian Journal of Veterinary Research, 56, 127-134.
- KWIECIEN, J. M., LITTLE, P. B. & HAYES, M. A., 1994. Adherence of Haemophilus somni to tumor necrosis factor-alpha-stimulated bovine endothelial cells in culture. Canadian Journal of Veterinary Research, 58, 211-219.
- LAMONT, H. H. & HUNT, B. W., 1982. Haemophilus somni and conjunctivitis. Veterinary Record, 117, 21.
- LANE, M. V. & LEDER, R. L., 1988. Unilateral nephrectomy for chronic Haemophilus somni-associated nephritis in a cow. Compendium on Continuing Education for the Practising Veterinarian, 10, 522-525.
- LARSON, R. L. & STEP, D. L., 2012. Evidence-based effectiveness of vaccination against Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni in feedlot cattle for mitigating the incidence and effect of bovine respiratory disease complex. Veterinary Clinics of North America: Food Animal Practice, 28(1), 97-106.
- LEDERER, J. A., BROWN, J. F. & CZUPRYNSKI, C. J., 1987. ‘Haemophilus somni’, a facultative intracellular pathogen of bovine mononuclear phagocytes. Infection and Immunity, 55, 381-387.
- LITTLE, P. B., 1986. Haemophilus somni complex: Pathogenesis of the septicaemic thrombotic meningoencephalitis. Canadian Veterinary Journal, 27, 94-96
- LITTLE, P. B. & SORENSEN, D. K., 1969. Bovine polioencephalomalacia, infectious embolic meningoencephalitis, and acute lead poisoning in feedlot cattle. Journal of the American Veterinary Medical Association, 155, 1892-1903.
- LO, K. L., KIMBALL, R. A., LEHMANN, J., GERSHWIN, L. J., WORBY, C. & CORBEIL, L. B., 2012. Antibody responses of calves to Histophilus somnirecombinant IbpA subunits. Comparative Immunology, Microbiology & Infectious Diseases, 35, 453-459.
- LOY, J. D., LEGER, L., WORKMAN, A. M., CLAWSON, M. L., BULUT, E. & WANG, B., 2018. Development of a multiplex real-time PCR assay using two thermocycling platforms for detection of major bacterial pathogens associated with bovine respiratory disease complex from clinical samples. Journal of Veterinary Diagnostic Investigation, 30, 837-847.
- MAGSTADT, D. R., SCHULER, A. M., COETZEE, J. F., KRULL, A. C., O'CONNOR, A. M., COOPER, V. L. & ENGELKEN, T. J., 2018. Treatment history and antimicrobial susceptibility results for Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni isolates from bovine respiratory disease cases submitted to the Iowa State University Veterinary Diagnostic Laboratory from 2013 to 2015. Journal of Veterinary Diagnostic Investigation, 30, 99-104.
- MARGINEDA, C. A., O'TOOLE, D., PRIETO, M., UZAL, F. A. & ZIELINSKI, G. C., 2019. Histophilus somnimyocarditis and leptomeningitis in feedlot cattle: case report and occurrence in South America. Journal of Veterinary Diagnostic Investigation, 31, 893-898.
- MARTIN, S. W., HARLAND, R. J., BATEMAN, K. G. & NAGY, E., 1998. The association of titers to Haemophilus somni, and other putative pathogens, with the occurrence of bovine respiratory disease and weight gain in feedlot calves. Canadian Journal of Veterinary Research, 62, 262-267.
- MCEWEN, S. A. & HULLAND, T. J., 1985. Haemophilus somni-induced otitis and meningitis in a heifer. Canadian Veterinary Journal, 26, 7-8.
- MCQUISTON, J. H., MCQUISTON, J. R., COX, A. D., WU, Y., BOYLE, S. M. & INZANA, T. J., 2000. Characterization of a DNA region containing 5’-(CAAT)n-3’ DNA sequences involved in lipooligosaccharide biosynthesis in Haemophilus somnus. Microbial Pathogenesis, 28, 301-312.
- METZ, A. L., HAGGARD, D. L. & MAKOMAKI, M. R., 1984. Chronic suppurative orchiepididymitis associated with Haemophilus somni in a calf. Journal of the American Veterinary Medical Association, 184, 1507-1508.
- MILLER, R. B., LEIN, D. H., MCENTEE, K. E., HALL, C. E. & SHIN, S., 1983. Histophilus somni infection of the reproductive tract of cattle: A review. Journal of the American Veterinary Medical Association, 182, 1390-1392.
- MILLER, R. B., VAN CAMP, S. D. & BARNUM, D. A., 1983. The effects of intra-amniotic inoculation of Haemophilus somni on the bovine fetus and dam. Veterinary Pathology, 20, 574-583.
- MOHD-ZAIN, Z., TURNER, S. L., CERDEÑO-TÁRRAGA, A. M., LILLEY, A. K., INZANA, T. J., DUNCAN, A. J., HARDING, R. M., HOOD, D. W., PETO, T. E. & CROOK, D. W., 2004. Transferable antibiotic resistance elements in Haemophilus influenzae share a common evolutionary origin with a diverse family of coherent genomic islands. Journal of Bacteriology, 86, 8114-8122.
- MOMOTANI, E., YABUKI, Y., MIHO, H., ISHIKAWA, Y. & YOSHINO, T., 1985. Histopathologic evaluation of disseminated intravascular coagulation in Haemophilus somni infection in cattle. Journal of Comparative Pathology, 95, 15-23.
- MORCK, D. W., MERRILL, J. K., THORLAKSON, B. E., OLSON, M. E., TONKINSON, L. V. & COSTERTON, J. W., 1993. Prophylactic efficacy of tilmicosin for bovine respiratory tract disease. Journal of the American Veterinary Medical Association, 202, 273-277
- MORTER, R. L. & AMSTUTZ, H. E., 1983. Evaluating the efficacy of a Haemophilus somni bacterin in a controlled field trial. The Bovine Practitioner, 18, 82-83.
- NATION, P. N., FRELIER, P. F., GIFFORD, G. A. & CARNAT, B. D., 1983. Otitis in feedlot cattle. Canadian Veterinary Journal, 24, 238
- NIVARD, J. L., WARD, G. E., STEPHENS, J. B. & MAHESWARAN, S. K., 1982. Model infection of the chicken embryo with Haemophilus somni. American Journal of Veterinary Research, 43, 1790-1792.
- O'TOOLE, D., ALLEN, T., HUNTER, R. & CORBEIL, L. B., 2009. Diagnostic exercise: Myocarditis due to Histophilus somni in feedlot and backgrounded cattle. Veterinary Pathology, 46,1015-1017.
- O'TOOLE, D. & SONDGEROTH, K. S., 2016. Histophilosis as a Natural Disease. Current Topics in Microbiology and Immunology, 396, 15-48.
- PAN, Y., FISHER, T., OLK, C. & INZANA, T. J., 2014. Detection of antibodies to the biofilm exopolysaccharide of Histophilus somni following infection in cattle by enzyme-linked immunosorbent assay. Clinical Vaccine Immunology, 21, 1463.
- PAN, Y., TAGAWA, Y., CHAMPION, A., SANDAL, I. & INZANA, T. J., 2018. Histophilus somni survives in bovine macrophages by interfering with phagosome-lysosome fusion but requires IbpA for optimal serum resistance. Infection and Immunity, 86,e00365-18.
- PANCIERA, R. J., DAHLGREN, R. R. & RINKER, H. B., 1968. Observations on septicaemia of cattle caused by a Haemophilus-like organism. Pathologia Veterinaria, 5, 212-226.
- PHILBEY, A. W., GLASTONBURY, J. R., ROTHWELL, J. T., LINKS, I. J. & SEARSON, J. E., 1991. Meningoencephalitis and other conditions associated with Histophilus ovis infection in sheep. Australian Veterinary Journal, 68, 387-390
- POTGIETER, L. N. D., HELMAN, R. G., GREENE, W., BREIDER, M. A., THURBER, E. T. & PEETZ, R. H., 1988. Experimental bovine respiratory tract disease with Haemophilus somni. Veterinary Pathology, 25, 124-130.
- PRESCOTT, J. F. & YIELDING, K. M., 1990. In vitro susceptibility of selected veterinary bacterial pathogens to ciprofloxacin, enrofloxacin and norfloxacin. Canadian Journal of Veterinary Research, 54, 195-197.
- PRITCHARD, D. G., SHREEVE, J. & BRADLEY, R., 1979. The experimental infection of calves with a British strain of Haemophilus somni. Research in Veterinary Science, 26, 7-11.
- RUBY, K. W., GRIFFITH, R. W., GERSHWIN, L. J. & KAEBERLE, M. L., 2000. Haemophilus somni-induced IgE in calves vaccinated with commercial monovalent H. somni bacterins.Veterinary Microbiology, 76, 373-383.
- RUBY, K. W., GRIFFITH, R. W. & KAEBERLE, M. L., 2002. Histamine production by Haemophilus somnus. Comparative Immunology and Microbiology, 25, 13-20.
- SALMON, S. A., WATTS, J. L. & YANCEY, R. J. J., 1996. In vitro activity of ceftiofur and its primary metabolite, desfuroylceftiofur, against organisms of veterinary importance. Journal of Veterinary Diagnostic Investigation, 8, 332-336.
- SANDAL, I. & INZANA, T. J., 2010. Histophilus somni Host-Pathogen Interactions: A Genomic Window into Virulence. Trends in Microbiology, 18, 90-99.
- SANDAL, I., INZANA, T. J., MOLINARO, A., DE CASTRO, C., SHAO, J. Q., APICELLA, M. A., COX, A. D., ST MICHAEL, F. & BERG, G., 2011. Identification, structure, and characterization of an exopolysaccharide produced by Histophilus somni during biofilm formation. BMC Microbiology, 11, 186.
- SANDAL, I., SHAO, J. Q., ANNADATA, S., APICELLA, M. A., BOYE, M., JENSEN, T. K., SAUNDERS, G. K. & INZANA, T. J., 2009. Histophilus somni biofilm formation in cardiopulmonary tissue of the bovine host following respiratory challenge. Microbes and Infection, 11, 254e263.
- SANDAL, I. W., HONG, W., SWORDS, W. E. & INZANA, T. J., 2007. Characterization and comparison of biofilm development by pathogenic and commensal isolates of Histophilus somni. Journal of Bacteriology, 189, 8179-8185.
- SAUER, K., CAMPER, A. K., EHRLICH, G. D., COSTERTON, J. W. & DAVIES, D. G., 2002. Pseudomonas aeruginosadisplays multiple phenotypes during development as a biofilm. Journal of Bacteriology. 184, 1140-1154.
- SAUNDERS, J. R. & JANZEN, E. D., 1980. Haemophilus somni infections. II. A Canadian field trial of a commercial bacterin: Clinical and serological results. Canadian Veterinary Journal, 21, 219-224.
- SAUNDERS, J. R., THIESSEN, W. A. & JANZEN, E. D., 1980. Haemophilus somni infections. I. A ten year (1969–1978) retrospective study of losses in cattle herds in western Canada. Canadian Veterinary Journal, 21, 119-123
- SCANLAN, C. M., 1988. Introduction to Veterinary Bacteriology, Ames, Iowa: Iowa State University Press.
- SCHIEFER, B., WARD, G. E. & MOFFAT, R. E., 1978. Correlation of microbial and histological findings in bovine fibrinous pneumonia. Veterinary Pathology, 15, 313-321.
- SIDDARAMAPPA, S., CHALLOCOMBE, J. F., DUNCAN, A. J., GILLASPY, A. F., CARSON, M., GIPSON, J., ORVIS, J., ZAITSHIK, J., BARNES, G., BRUCE, D., CHERTKOV, O., DETTER, J. C., HAN, C. S., TAPIA, R., THOMPSON, L. S., DYER, D. W. & INZANA, T. J., 2011. Horizontal gene transfer in Histophilus somni and its role in the evolution of pathogenic strain 2336, as determined by comparative genomic analyses. BMC Genomics, 12, 570.
- SIDDARAMAPPA, S. & INZANA, T. J., 2004. Haemophilus somnus virulence factors and resistance to host immunity. Animal Health Research Reviews, 5, 79-94.
- SLEE, K. J. & STEPHENS, L. R., 1985. Selective medium for isolation of Haemophilus somni from cattle and sheep. Veterinary Record, 116, 215-217.
- STAROST, M. F., 2001. Haemophilus somni isolated from a urachal abscess in a calf. Veterinary Pathology, 38, 547-548.
- STEPHENS, L. R., 1990. Positive and negative aspects of host immune response to Haemophilus, Actinobacillus and Pasteurella. Canadian Journal of Veterinary Research, 54 Suppl, S41-S44
- STEPHENS, L. R., HUMPHREY, J. D., LITTLE, P. B. & BARNUM, D. A., 1983. Morphological, biochemical, antigenic, and cytochemical relationships among Haemophilus somni, Haemophilus agni, Haemophilus haemoglobinophilus, Histophilus ovis and Actinobacillus seminis. Journal of Clinical Microbiology, 17, 728-737
- STEPHENS, L. R., LITTLE, P. B., WILKIE, B. N. & BARNUM, D. A., 1981. Infectious thromboembolic meningoencephalitis in cattle: A review. Journal of the American Veterinary Medical Association, 178, 378-384.
- STEPHENS, L. R., SLEE, K. J., POULTON, P., LARCOMBE, M. & KOSIOR, E., 1986. Investigation of purulent vaginal discharge in cows, with particular reference to Haemophilus somni. Australian Veterinary Journal, 63, 182-185.
- STUART, F. A., CORBEL, M. J., RICHARDSON, C., BREWER, R. A., BRADLEY, R. & BRIDGES, A. W., 1990. Experimental Haemophilus somni infection in pregnant cattle. British Veterinary Journal, 146, 57-67.
- SYLTE, M. J., CORBEIL, L. B., INZANA, T. J. & CZUPRYNSKI, C. J., 2001. Haemophilus somni induces apoptosis in bovine endothelial cells in vitro. Infection and Immunity, 69, 1650-1660.
- TAGAWA, Y., SANDERS, J. D., UCHIDA, I., BASTIDA-CORCUERA, F. D., KAWASHIMA, K. & CORBEIL, L. B., 2005. Genetic and functional analysis of Haemophilus somnushigh molecular weight-immunoglobulin binding proteins. Microbial Pathogenesis, 39, 159-170.
- TAYLOR, J. D., FULTON, R. W., LEHENBAUER, T. W., STEP, D. L. & CONFER, A. W., 2010a. The epidemiology of bovine respiratory disease: What is the evidence for predisposing factors? Canadian Veterinary Journal, 51, 1095-1102.
- TAYLOR, J. D., FULTON, R. W., LEHENBAUER, T. W., STEP, D. L. & CONFER, A. W., 2010b. The epidemiology of bovine respiratory disease: what is the evidence for preventive measures? Canadian Veterinary Journal, 51, 1351-1359.
- TEGTMEIER, C., ANGEN, Ø. & AHRENS, P., 2000. Comparison of bacterial cultivation, PCR, in situ hybridization and immunohistochemistry as tools for diagnosis of Haemophilus somni pneumonia in cattle. Veterinary Microbiology, 76, 385-394.
- TEGTMEIER, C., ANGEN, Ø., GRELL, S. N., RIBER, U. & FRIIS, N. F., 2000. Aerosol challenge of calves with Haemophilus somni and Mycoplasma dispar. Veterinary Microbiology, 72, 229-239.
- TEGTMEIER, C., JENSEN, N. E. & JENSEN, H. E., 1995. Development of a peroxidase-antiperoxidase (PAP) technique for the identification of Haemophilus somni in pneumonic calf lungs in Denmark. Acta Pathologica, Microbiologica et Immunologica Scandinavica, 103, 540-547.
- TEGTMEIER, C., UTTENTHAL, A., FRIIS, N. F., JENSEN, N. E. & JENSEN, H. E., 1999. Pathological and microbiological studies on pneumonic lungs from Danish calves. Journal of Veterinary Medicine Series B, 46, 693-700.
- THANTHRIGE-DON, N., LUNG, O. O., FURUKAWA-STOFFER, T., BUCHANAN, C., JOSEPH, T., GODSON, D. L., GILLEARD, J., ALEXANDER, T. & AMBAGALA, A. P., 2018. A novel multiplex PCR-electronic microarray assay for rapid and simultaneous detection of bovine respiratory and enteric pathogens. Journal of Virological Methods, 261, 51-62.
- THOMAS, A. C., BAILEY, M., LEE, M. R. F., MEAD, A., MORALES-AZA, B., REYNLDS, R., VIPOND, B., FINN, A. & EISLER, M. C., 2019. Insights into Pasteurellaceae carriage dynamics in the nasal passages of healthy beef calves. Scientific Reports 9, 11943.
- THOMPSON, K. G. & LITTLE, P. B., 1981. Effect of Histhophilus somni on bovine endothelial cells in organ culture. American Journal of Veterinary Research, 42, 748-754.
- THOMPSON, K. G., VICKERS, M. C., STEVENSON, B. J. & DAVIDSON, G. W., 1987. Thromboembolic meningoencephalitis caused by Haemophilus somni in a bull calf — a new disease in New Zealand. New Zealand Veterinary Journal, 35, 5-7.
- THOMSON, M. S., LAUERMANN, L. H. & WILT, G. R., 1990. Monoclonal antibody in the identification of Haemophilus somni. Journal of Veterinary Diagnostic Investigation, 2, 116-119.
- THOMSON, M. S., STRINGFELLOW, D. A. & LAUERMAN, L. H., 1988. Adherence of Haemophilus somni to bovine embryos after in vitro exposure. American Journal of Veterinary Research, 49, 63-66.
- UENO, Y., TERATANI, C., MISUMI, W., HOSHINOO, K., TAKAMATSU, D., TAGAWA, Y. & KATSUDA, K., 2018. A predominant clonal thromboembolic meningoencephalitis group of Histophilus somniassigned by major outer membrane protein gene sequencing and pulsed-field gel electrophoresis. Frontiers in Veterinary Science, 5, 221.
- VAN DONKERSGOED, J., GUENTHER, C., EVANS, B. N., POTTER, A. A. & HARLAND, R. J., 1995. Effects of various vaccination protocols on passive and active immunity to Pasteurella haemolytica and Haemophilus somnus in beef calves. Canadian Veterinary Journal, 36, 424-429.
- VAN DONKERSGOED, J., JANZEN, E. D. & HARLAND, R. J., 1990. Epidemiological features of calf mortality due to hemophilosis in a large feedlot. Canadian Veterinary Journal, 31, 821-825
- VAN DONKERSGOED, J., JANZEN, E. D., POTTER, A. A. & HARLAND, R. J., 1994. The occurrence of Haemophilus somni in feedlot calves and its control by postarrival prophylactic mass medication. Canadian Veterinary Journal, 35, 573-580.
- VAN DONKERSGOED, J., POTTER, A. A., MOLLISON, B. & HARLAND, R. J., 1994. The effect of a combined Pasteurella haemolytica and Haemophilus somnus vaccine and a modified-live bovine respiratory syncytial virus vaccine against enzootic pneumonia in young beef calves. Canadian Veterinary Journal, 35, 239-241.
- VAN DREUMEL, A. A. & KIERSTEAD, M., 1975. Abortion associated with Haemophilus somni infection in a bovine fetus. Canadian Veterinary Journal, 16, 367-370.
- VOGEL, G., NICOLET, J., MARTIG, J., TSCHUDI, P. & MEYLAN, M., 2001. Pneumonia in calves: Characterization of the bacterial spectrum and the resistance patterns to antimicrobial drugs. Schweizer Archiv für Tierheilkunde, 143, 341-350.
- WARD, A. C., DYER, N. W. & CORBEIL, L. B., 1999. Characterization of putative Haemophilus somni isolates from tonsils of American bison (Bison bison). Canadian Journal of Veterinary Research, 63, 166-169.
- WATTS, J. L., YANCEY, R. J. J., SALMON, S. A. & CASE, C. A., 1994. A 4-year survey of antimicrobial susceptibility trends for isolates from cattle with bovine respiratory disease in North America. Journal of Clinical Microbiology, 32, 725-731.
- WHITELEY, M., BANGERA, M. G., BUMGARNER, R. E., PARSEK, M. R., TEITZEL, G. M., LORY, S. & GREENBERG, E. P., 2001. Gene expression in Pseudomonas aeruginosabiofilms. Nature, 413, 860-864.
- WIDDERS, P. R., PAISLY, I. G., GOGOLEWSKI, R. P., EVERMANN, J. F., SMITH, J. W. & CORBEIL, I. B., 1986. Experimental abortion and the systemic immune response to Haemophilus somni in cattle. Infection and Immunity, 54, 555-560
- WILLIAMS, J. M., SMITH, G. L. & MURDOCK, F. M., 1978. Immunogenicity of a Haemophilus somni bacterin in cattle. American Journal of Veterinary Research, 39, 1756-1762.
- WISSELINK, H. J., CORNELISSEN, J., VAN DER WAL, F. J., KOOI, E. A., KOENE, M. G., BOSSERS, A., SMID, B., DE BREE, F. M. & ANTONIS, A. F. G., 2017. Evaluation of a multiplex real-time PCR for detection of four bacterial agents commonly associated with bovine respiratory disease in bronchoalveolar lavage fluid. BMC Veterinary Research, 13, 221.
- WORBY, C. A., MATTOO, S., KRUGER, R. P., CORBEIL, L. B., KOLLER, A., MENDEZ, J. C., ZEKARIAS, B., LAZAR, C. & DIXON, J. E., 2009. The fic domain: regulation of cell signaling by adenylylation. Molecular Cell, 34, 93-103.
- YANG, Y. F., SYLTE, M. J. & CZUPRYNSKI, C. J., 1998. Apoptosis: A possible tactic of Haemophilus somni for evasion of killing by bovine neutrophils? Microbial Pathogenesis, 24, 351-359.
- YARNALL, M. & CORBEIL, L. B., 1989. Antibody response to Haemophilus somnusFc receptor. Journal of Clinical Microbiology, 27, 111-117.
- YEO, H. J., YOKOYAMA, T., WALKIEWICZ, K., KIM, Y., GRASS, S. & GEME, J. W. R., 2007. The structure of the Haemophilus influenzaeHMW1 pro-piece reveals a structural domain essential for bacterial two-partner secretion. Journal of Biological Chemistry, 282, 31076-31084.
- ZEKARIAS, B., MATTOO, S., WORBY, C., LEHMANN, J., ROSENBUSCH, R. F. & CORBEIL, L. B., 2010. Histophilus somni IbpA DR2/Fic in virulence and immunoprotection at the natural host alveolar epithelial barrier. Infection and Immunity, 78, 1850-1858.
- ZEKARIAS, B., O'TOOLE, D., LEHMANN, J. & CORBEIL, L. B., 2011. Histophilus somniIbpA Fic cytotoxin is conserved in disease strains and most carrier strains from cattle, sheep and bison. Veterinary Microbiology, 149, 177-185.
- ZHANG, M., HILL, J. E., GODSON, D. L., NGELEKA, M., FERNANDO, C. & HUANG, Y., 2019. The pulmonary virome, bacteriological and histopathological findings in bovine respiratory disease from western Canada. Transboundary Emerging Diseases, doi:10.1111/tbed.13419.




