- Infectious Diseases of Livestock
- Part 3
- Tuberculosis
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Tuberculosis
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Tuberculosis
D V COUSINS, H F K A HUCHZERMEYER, J F T GRIFFIN, G K BRU¨CKNER, I B J VAN RENSBURG AND N P J KRIEK
Mycobacterium bovis infection in cattle
Synonyms: Bovine tuberculosis, tuberkulose by beeste (Afrik.)
Bovine tuberculosis existed in the Mediterranean littoral before classical times. It spread from northern Italy to western Europe and Great Britain and from there infected cattle carried the disease to many parts of the world that had been colonized.105, 129 Currently, the disease occurs globally and it appears to be an ever-increasing problem. Even in developed countries, major difficulties are experienced to control and eradicate the disease from cattle. The distribution of the disease and its prevalence in the various countries can be obtained from sources such as World Animal Health compiled by the Office International des Epizooties (OIE) on an ongoing basis,5 and others.27 Bovine tuberculosis is an important zoonosis, its prevalence in humans showing a close correlation with the level of infection in the cattle population of a specific country.27
In recent years it has become evident that infection with Mycobacterium bovis, the cause of the infection in cattle, is common in a wide variety of wildlife in various parts of the world. The infection in wildlife is important not only from the perspective of the value of some species of wildlife, some of which are endangered, but because of the role that wildlife maintenance hosts of the disease can play in sustaining the infection in domestic stock.33 The complex nature of the epidemiology of bovine tuberculosis, and the role of wildlife in sustaining the infection, are becoming clearer as time goes on and have an adverse effect on the efficacy of control measures that are currently in use in developed and developing countries.
The first reference to the occurrence of tuberculosis in animals in South Africa was made by Hutcheon in 188068 and it appears unlikely that bovine tuberculosis existed in South Africa prior to the importation of European breeds of cattle towards the end of the nineteenth century.62 In 1902 it was reported that cattle in Natal were free of tuberculosis but that many animals imported from Australia, the Argentine, Madagascar and England, that reacted to the tuberculin test, were destroyed upon arrival in Durban.
Subsequently, the prevalence of the disease in slaughtered cattle remained low in South Africa between 1917 and 1929. Its spread in southern Africa was originally confined to dairy herds but since the early 1970s beef herds have been found to be infected.
Since bovine tuberculosis was first recorded in Zimbabwe in 1908 in a heifer imported from the Western Cape Province, South Africa, only sporadic cases have been diagnosed. A national testing scheme for tuberculosis was instituted from 1950 to 1978. The prevalence remained very low, with one or none of the slaughtered animals per year showing tuberculous lesions.61
Namibia is considered to be virtually free of tuberculosis in livestock, and all diagnosed cases can be traced back to imported animals that originated mainly from South Africa and Germany.128 From 1913 to 1984, of 66 888 head of cattle tested, 163 reacted positive to the tuberculin test and 284 gave suspicious reactions. At abattoirs only seven animals were found to be tuberculous.2
Tuberculosis in animals in Swaziland is rarely diagnosed. Its prevalence in cattle is estimated to be below 0,5 per cent and it has not been diagnosed in wild animals.37
Epidemiology
Tuberculosis in cattle is caused predominantly by M. bovis, of which cattle are the maintenance hosts. In New Zealand the possum (Trichosurus vulpecula) and fallow deer (Dama dama), in England the badger (Meles meles), in some European countries swine and deer, and in the USA and Canada bison (Bison bison) and elk (Cervus elaphus) are additional maintenance hosts. In the USA, white-tailed deer (Odocoileus virginianus) have also become infected and are now considered a maintenance host for M. bovis.
Mycobacterium bovis infection has recently been diagnosed in African buffalo (Syncerus caffer), which is considered to be a maintenance host, and various other species of wildlife in some game parks and commercial game ranches in South Africa. Tuberculosis in the water buffalo (Bubalus bubalis) in some countries and free-living African buffaloes in African game reserves may therefore constitute a source of infection when they mingle with livestock on the borders of conservation areas.33 Kudu (Tragelaphus strepsiceros) in certain areas in South Africa may play a limited role in the transmission of M. bovis to cattle.139
Infection with Mycobacterium tuberculosis in cattle is rare, but it may cause reactions in tuberculin-tested cattle. Infection with members of the Mycobacterium avium intracellulare complex (MAIC) is not a major disease problem in cattle but can confound tuberculosis eradication programmes in cattle. Cattle infected with M. avium become sensitized to mammalian tuberculin, which may cause false positive results in the intradermal tuberculin test. In cattle the sensitization to M. avium and M. tuberculosis infections may disappear soon after they are removed from contact with either infected birds or humans infected with M. tuberculosis. Infection and disease due to M. bovis in other susceptible species in contact with cattle, including humans, decline dramatically when comprehensive eradication programmes in cattle are in progress or have been successfully concluded.20 However, should M. bovis infection become established in a susceptible wildlife population, such as African buffalo, total eradication of the disease from cattle becomes difficult if not impossible.33
Humans with open tuberculosis caused by M. bovis can infect cattle by the aerogenous route when handling them or by contaminating the closed environment of a stable by spitting or urinating.22, 102, 122, 146, 147
Mycobacterium tuberculosis infection in cattle usually does not result in progressive disease, though localized lesions do develop. Infected cattle are generally not contagious, although excretion of bacilli in animals that are not manifesting signs of the infection has been reported. Bacilli may persist for some time in the organs and tissues. In one investigation it was found that most cattle experimentally infected with M. tuberculosis reacted negatively to the tuberculin test after some months.154
The primary mode of spread of bovine tuberculosis between herds is by the introduction of infected animals into non-infected herds. In contrast to European countries, close contact in confined spaces is not a prerequisite for the spread of the disease. Infections have been found in extensive cattle-breeding areas where ranching conditions prevail and in which there is little or no close contact between animals in confined spaces for prolonged periods. Under similar conditions in Australia, tuberculosis in extensively kept cattle is thought to be due to their habit of congregating daily in large numbers at watering points under very dusty conditions.93
Although tuberculosis in cattle has been recognized for a long time, many features of its epidemiology have not been elucidated. The lack of knowledge is such that even in some developed countries, such as the UK and Ireland, it has not been possible to eradicate the disease in spite of intensive efforts and investment of large sums of money over decades. The most important factors determining the occurrence and spread of tuberculosis in a herd of cattle are the herd size, number of infected individuals, the number of young stock exposed to infected animals, existing farm practices, and measures taken to prevent spread of the disease. The mode of transmission in cattle is mainly horizontal but not all infected animals transmit the disease. Most cattle suffering from pulmonary tuberculosis and tuberculous mastitis are, however, infectious, and in some infected individuals urine, vaginal secretions, semen or faeces may also contain tubercle bacilli and act as a means of transmitting the disease between animals.75 The risk of infection from infected cattle is dependent on them shedding the organism. Bacteria are shed consistently soon after the initial infection has occurred; at a later stage, shedding becomes intermittent. Shedding of mycobacteria has also been detected in animals that respond negatively to the tuberculin test106 and the likelihood of this happening is increased by the practice of introducing animals from negative cohorts from infected herds.15 Cattle from which M. bovis cannot be cultured should not automatically be disregarded as a potential source of infection.106
Possible routes of infection with M. bovis are respiratory, alimentary, congenital, cutaneous, venereal and via the teat canal. In practice, infection in intensively managed animals is acquired almost exclusively aerogenously by the inhalation of infected droplets from a coughing or sneezing animal with open pulmonary tuberculosis, or of infected dust particles. 48, 76 In stables and barns housing tuberculous cattle such droplets and particles may be constantly present in the air.69 Oral and alimentary tract infection may be primary in calves drinking the milk of cows with tuberculous mastitis,91 or it may be secondary from swallowing mycobacterialaden exudate expectorated from the lungs. Tonsillar infection with secondary involvement of regional lymph nodes may result from the ingestion of bacilli in the food or in neonatal animals through grooming and mucosal spread. More recently it has been suggested that the primary infection may occur in the tonsils and that this upper respiratory infection may be another source of bacteria that may be shed in respiratory secretions.106
The congenital route is important in chronically infected cattle herds with a high prevalence of tuberculosis because of the relatively high prevalence of tuberculous endometritis (some authors consider this to be as high as 5 per cent). In contrast, in cattle herds subjected to routine tuberculin test surveillance, limited lesions that are no more extensive than a primary thoracic complex, are the extreme presentation seen.
Infection via damaged skin is possible. For example, in humans it might happen in meat inspectors who have cut themselves with a knife contaminated with mycobacteria while examining tuberculous cattle, or in a laboratory worker who has pricked him- or herself with a Pasteur pipette containing a mycobacterial suspension. There is a recent documented case of tuberculosis in a veterinarian who suffered a cut during the course of a necropsy of a tuberculous possum.25 Percutaneous infection as a result of scratching the ears with contaminated hind legs has been described in kudus in the Eastern Cape Province in South Africa,115, 139 and through the bites of badgers in England.100 Venereal transmission is rare, but has been encountered in cattle.136 Infusion of contaminated material via the teat canal may result in tuberculous mastitis.
Although one can only speculate on the period of infectiousness, it might be extremely long if no herd surveillance scheme is in operation. An undiagnosed carrier that excretes bacteria may transmit the infection for months or even years. Large numbers of tubercle bacilli may be excreted in the milk of cows with tuberculous mastitis; there may be sufficient numbers of viable bacilli in the milk of a cow to contaminate the milk of 100 uninfected cows when such milk is pooled for transportation. Separated, defatted milk purchased by farmers from milk-processing factories to feed their calves can in this way greatly contribute to the spread of M. bovis.
The feeding of uncooked infected carcasses to animals is a potential source of infection; affected lymph nodes, in particular, being important in this respect. Muscular tissue (meat) is usually free of tubercle bacilli.48 Diseased tissues of cattle generally do not contain as many living tubercle bacilli as does the sputum of humans with open lung lesions.
In South Africa, tuberculosis caused by M. bovis has been diagnosed in wild antelope, namely kudu and Cape duiker (Sylvicapra grimmia) in the Albany district of the Eastern Cape Province,109, 139 and African buffalo in the Kruger National Park,9 and in some of the game parks (Umfolozi and Hluhluwe) in KwaZulu-Natal Province that are under the jurisdiction of the KwaZulu-Natal Parks Board.45 In some of these areas the infection has subsequently spread to at least 14 other mammalian species, including lion (Panthera leo), leopard (Panthera pardus), hyena (Crocuta crocuta), cheetah (Acinonyx jubatus), baboon (Papio ursinus), honey badger (Mellivora capensis), and warthog (Phacochoerus aethiopicus); these are considered to be spill-over species. In other parts of Africa tuberculosis in free-living wild animals has been reported in lechwe (Kobus leche) in Zambia,14, 69 and in African buffalo in Uganda.57, 152, 153 In countries on other continents tuberculosis caused by M. bovis has been reported to occur in the following free-living wild animals: the possum in New Zealand,40 the badger in England,17, 63 wild boar in Spain,7 and sika (Cervus nippon) and fallow deer in California.80 Tuberculosis has been described in camels (Camelus dromedarius) and farmed water buffalo in Egypt,41, 42 farmed fallow deer in New Zealand and Sweden, 34, 54, 65 red deer (Cervus elaphus) in England130 and Hungary,157 farmed elk in Canada43 and the USA,138 freeranging bison in Canada132 and the USA, and white-tailed deer in the USA.124
In captive wild animals, M. bovis, M. avium and M. tuberculosis (human strains) have all been responsible for numerous outbreaks in zoological gardens, involving many species ranging from non-human primates to elephants.103 Tuberculosis caused by a variant of M. tuberculosis has also been reported in both captive and wild pinnipeds in Australia29, 30 and South America.11
Humans are as susceptible to M. bovis as they are to M. tuberculosis. Once a tuberculous lesion develops in an organ, the appearance and the course of the disease are the same irrespective of whether it is caused by M. bovis or M. tuberculosis.102, 123 In humans, M. bovis infection often results in localized lesions and is most frequently the cause of cervical lymphadenitis (scrofula) and other forms of extrapulmonary tuberculosis in infancy and childhood. Although pulmonary tuberculosis due to M. bovis infection in humans is rare, the proportion of cases with pulmonary disease in developed nations has increased compared to extrapulmonary disease.28, 52, 150 Compared to the extent to which humans infect humans with tuberculosis, transmission from animals to humans is currently a minor problem. Extrapulmonary tuberculosis is typically transmitted via cows’ milk. Compulsory pasteurization was first introduced in 1898, not to protect humans, but to prevent tuberculosis in pigs and calves fed on separated milk obtained from infected herds. The varying use of pasteurization and inability to control the disease in cattle will in all likelihood have a significant effect on the prevalence of the disease in humans in developing countries.27
Pathogenesis
Some mycobacterial infections remain localized while others cause widely disseminated lesions. In the majority of cases very small lesions occur and the infection does not spread. For this reason many tuberculous infections cause very few clinical signs, if any, while in others, because of the varying location of the lesions, widely differing clinical signs are elicited. In general, and depending on the route of infection, primary foci of tuberculosis occur either in the gastrointestinal or in the respiratory tract. In the former site, lesions are found primarily in the lymphoid tissues, whereas in the respiratory system, both the lung parenchyma and the regional lymph nodes are affected. These variations are the consequence of unexplained variations in the pathogenicity of the organisms in different species, and the inherent characteristics of the organisms. Thus it appears from experimental results that the age of the animal, route of infection, strain of the organism, and the dose of bacteria will influence the length of time required for the disease to develop. 134 Lesions in different sites in the same animal may evolve independently of one another. At any given time some may be regressing, while others may be stable or expanding. 32
Many aspects of the pathogenesis of tuberculosis remain unknown even in humans and rodents, in spite of the fact that they are the focus of most of the current studies. The pathogenesis of the disease in cattle is less clear and one should refrain from extrapolating findings to other species because of the known differences in manifestation and pathogenesis of the disease in the various species. In several species the effects of a highly specific immune response results in a spectrum of lesions ranging from self-limiting infections to a life-threatening systemic disease.114 At every stage during the course of the tuberculous process the character of the lesions depends on the fluctuation between cellmediated immunity, associated with protective responses, and delayed type hypersensitivity that causes necrotising spreading lesions. In addition, the development of either an exudative or a proliferative response appears to depend on the concentration of bacterial antigen. Low concentrations of bacteria cause proliferative lesions, whereas high concentrations result in an exudative response.32 Some of the opportunistic mycobacteria which can be isolated from soil, dust and plants are able to cause persistent and slowly progressive infections which do not generate immune responses in the host.32
The outcome of a mycobacterial infection is dependent on whether an animal has been previously sensitized or not.148 The lesions in infected cattle may remain dormant for extended periods, progress or regress.106 In unsensitized hosts initial multiplication of the bacteria in macrophages results in a primary lesion, spread to the regional lymph node, and the development of a mild bacteraemia as soon as 20 days after the infection has taken place. This bacteraemia may terminate in the development of metastatic foci (a process referred to as post-primary dissemination). Following the development of cell-mediated immunity, however, most of the foci of infection, with the exception of those in certain ‘vulnerable sites’ such as the lungs, bone, kidneys and meninges, are destroyed. In the ‘vulnerable sites’ the bacterial population is reduced but persists and is not totally eliminated.148 The genotype of individual animals may also influence the response to a mycobacterial infection. Such examples exist in African zebu cattle that appear to be resistant to bovine tuberculosis, while introduced exotic breeds seem relatively susceptible. 14
In aerogenous infections, bacteria are inhaled in droplet nuclei. Only those droplets of 2 to 5 μmin diameter have the ability to reach the alveolar spaces of the lungs where they are deposited and the bacteria ingested by macrophages. Larger particles (or nuclei) are trapped in the upper airways and are removed by means of the mucocilliary apparatus, after which they are swallowed. The mucosal surfaces of the upper respiratory tract and the intestine are only infected after exposure to very large infective doses of bacteria.32
The initial elimination of mycobacteria depends on the microbicidal activities of the infected macrophages. When organisms initially multiply within the phagolysosomes of unactivated macrophages, their presence does not result in the death of many of the cells. When infected macrophages do die, the released bacteria are again phagocytosed by adjacent macrophages.
The ability of macrophages to kill mycobacteria is enhanced only after the development of a cellmediated immune response involving activated T cells (TH1). These cells produce cytokines, especially interferon and interleukin-12 (IL-12),55 which upregulate the microbiocidal properties of macrophages. The rapid development of the immune response after infection inhibits the growth of mycobacteria in lesions while they are still very small. The immune response that is elicited is complex and comprises a spectrum of immune responses of which the cell-mediated responses predominate; antibody responses only follow at a later stage.111 Various parameters of the cellmediated response develop at different stages of the disease.111
Macrophages at the site of infection are activated by direct contact with mycobacteria under the influence of lymphokines. The debris of necrotic macrophages sensitizes local lymphocytes. After sensitization, which may be as soon as two weeks after the initial infection, lymphocytes release lymphokines that attract, activate and increase the number of mononuclear cells at the site of infection, and contribute to cell death with the subsequent development of caseous necrosis.32, 135 During the early stages of the development of tubercles (tuberculous or mycobacterial granulomas), the round cell component of the reaction contain CD2+ T and WC1+ γδ lymphocytes, and B lymphocytes.16 The role of these cells in the development of the lesion in cattle remains unclear. Macrophages also play an integral part in the processing of mycobacterial antigens and the initiation of the immune response. Cell death caused by the delayed hypersensitivity reaction may be the result of lymphokines, toxic components of the bacteria, ischaemia, antigen-antibody reactions, hydrolytic enzymes and reactive oxygen intermediates released from apoptotic or necrotic macrophages and neutrophils, and tumour necrosis factor.32
The further development of mycobacterial granulomas appears to be dependent on a number of factors, the main ones being the ability of the mycobacteria to survive intracellularly, and their ability to induce a host response (delayed type hypersensitivity). Neutrophils initially enter into the development of the granuloma but they are soon replaced by macrophages. Certain glycolipid extracts (cord factor) from the bacteria inhibit chemotaxis and are leukotoxic, whereas others promote the intracellular survival of the bacteria in macrophages by inhibiting phagolysosome formation. At the early stages of development, tuberculous granulomas are composed of sensitized leukocytes and epithelioid cells. At this stage there is a dynamic engulfment of bacteria and necrosis of macrophages at the site of infection. As the development of the lesion progresses, the necrotic mass may become extensive. Langerhans’ giant cells appear in the granulomatous response and it also becomes surrounded by a zone of lymphocytes and other mononuclear cells. Often there is proliferation of connective tissue at the periphery which tends to encapsulate the lesion.134
The fate of focal tubercles is determined by the activity of macrophages within the lesion. When these cells have developed microbiocidal activities, they prevent expansion of the lesion; however, continuing intracellular bacterial growth enables the lesions to expand and eventually leads to haematogenous dissemination. This is usually the case in immature animals and in those that are immunodeficient.32 The multiplication of bacilli tends to be inhibited in lesions containing caseous exudate but if the exudate becomes liquefied their multiplication may be profuse. In the latter case, because of the high concentration of tuberculin-like products, the effect of the host’s cell-mediated immune response is severely detrimental and results in further local caseous necrosis. Bacteria may remain viable within the caseous centres of the larger tubercles for months or years and may become reactivated if the resistance of the host decreases.32
Caseous lesions may undergo liquefaction because of the effects of hydrolytic enzymes from macrophages and neutrophils. 134 This liquefied material is an excellent culture medium for the bacilli and contains large numbers of them.32 Liquefaction of the caseous exudate may lead to the disintegration of surrounding tissue, including the walls of ducts or tubes. If this occurs in the lungs, for instance, the entrance and spread of mycobacteria in the bronchial tree may result in extensive tubercular pneumonia.
Haematogenous spread may result either in miliary tuberculosis which is the result of a sudden massive bacteraemia and is manifested by the development of multiple small (millet-sized) lesions in many organs and tissues or, when it is more protracted and smaller numbers of bacteria enter the blood stream over a longer period, in the development of fewer, and larger, metastatic lesions which vary in age and in size. Live, infected macrophages may also enter blood and lymph vessels, ducts or body cavities and so they may disseminate the infection.134
Clinical signs and pathology
Bovine tuberculosis generally has a chronic, variable, and often subclinical course. It usually takes many months or even years before clinical signs develop.62 In most infected cattle the disease is inapparent, its presence only being detectable by the application of the tuberculin test, but if clinical signs are manifested, their nature depends on the organ system or systems involved and the severity of the infection.
Because of the complex nature of the disease, the clinical and pathological manifestations can vary substantially. The typical manifestation of the disease is also affected by the disease control measures that are applied: in countries where control is active and aggressive, the disease is primarily respiratory in nature and the extent of the lesions is limited. In those countries where the disease is not actively controlled, advanced disease and generalization are common. 106 Remissions and relapses, and in some instances, arrest and self-cure, may occur. Tuberculosis that involves the lymph nodes and to a lesser extent the lungs are the most common forms in cattle, while tuberculosis of the kidneys, liver, spleen, bone, joints, mammary gland, testes, uterus and ovaries is rare and is usually the consequence of haematogenous spread.
The distribution of lesions varies according to the route of infection, which may be by inhalation, per os, genital, via the teat canal, percutaneous, or congenital via the umbilical cord. Irrespective of the route of infection, complete or incomplete primary complexes are established.
A complete primary complex is one in which a lesion is present at the initial site of infection as well as in the regional lymph node. In an incomplete primary complex a lesion is found only in the lymph node regional to the site of infection.
Animals with disseminated lesions gradually become emaciated and anorexic and manifest a fluctuating temperature, have a dull hair coat, and may eventually become lethargic, but their eyes remain clear. Signs become more evident during stress such as that which may occur during the immediate post-calving period or as the result of malnutrition.
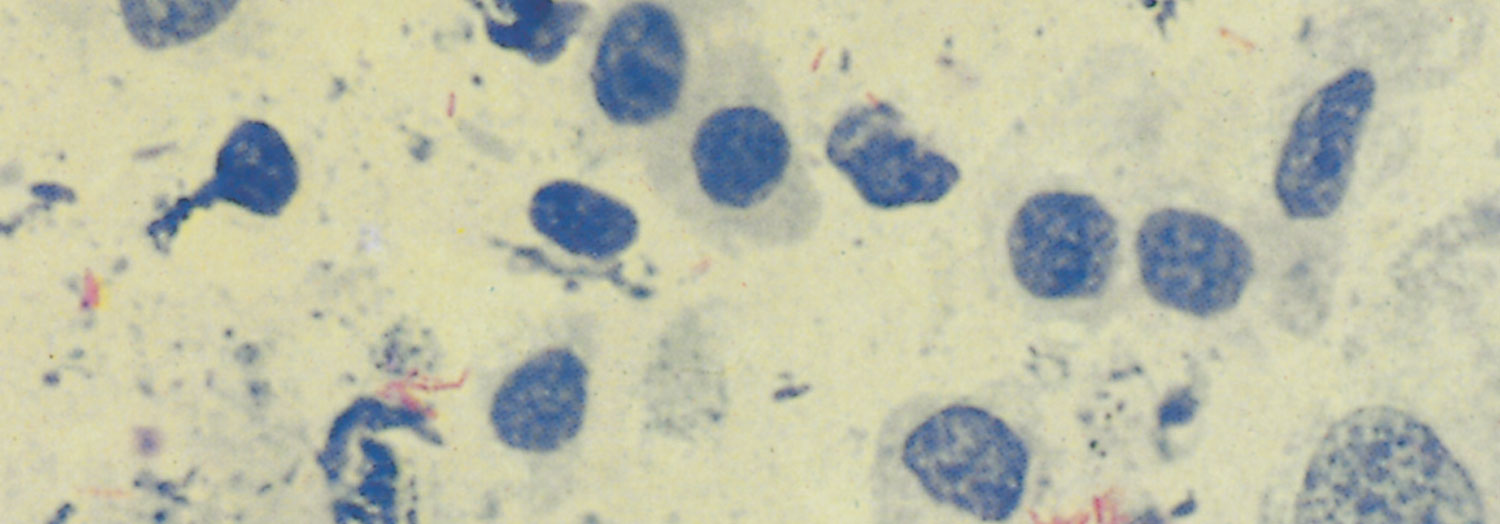
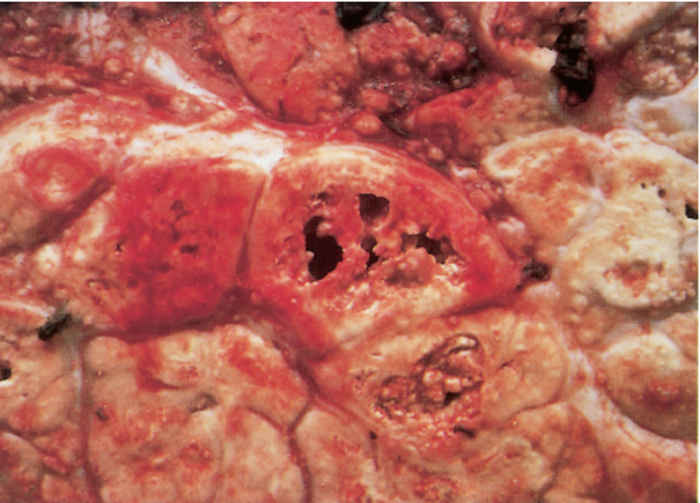
The classic lesion of tuberculosis, the tubercle, is a typical granuloma containing a central core of caseous necrotic tissue, part of which rapidly undergoes dystrophic calcification. This central mass is surrounded by epithelioid cells and Langhans’ giant cells, and more peripherally by lymphocytes, macrophages and varying degrees of fibroplasia; encapsulation may be complete or incomplete (Figure 196.1). Application of the Ziehl-Neelsen staining technique to sections or smears of affected tissue reveals varying numbers of pleomorphic, intracellular, acid-fast bacilli (Figure 196.2). Mycobacteria are Gram-positive and fluoresce after being stained with auramine. Exceptionally, the inflammatory reaction may be florid and markedly exudative, in which case large amounts of fibrin, haemorrhage and oedema occur in addition to the presence of macrophages and developing giant cells.
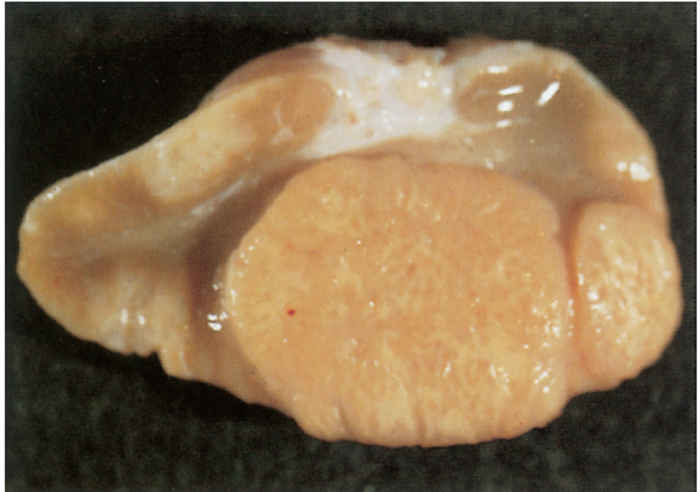
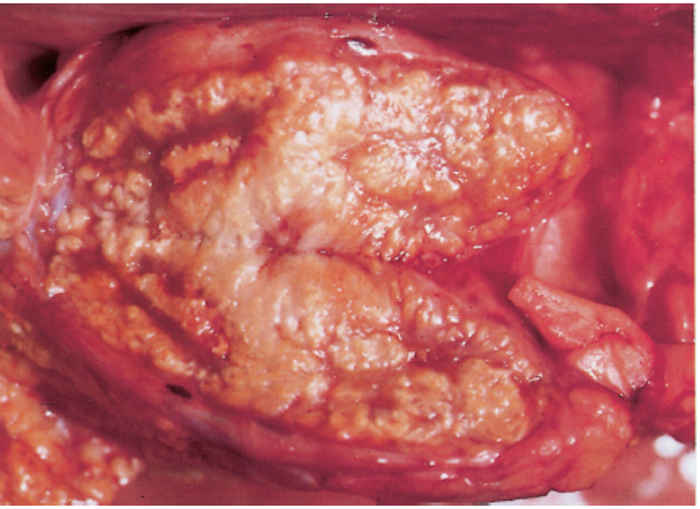
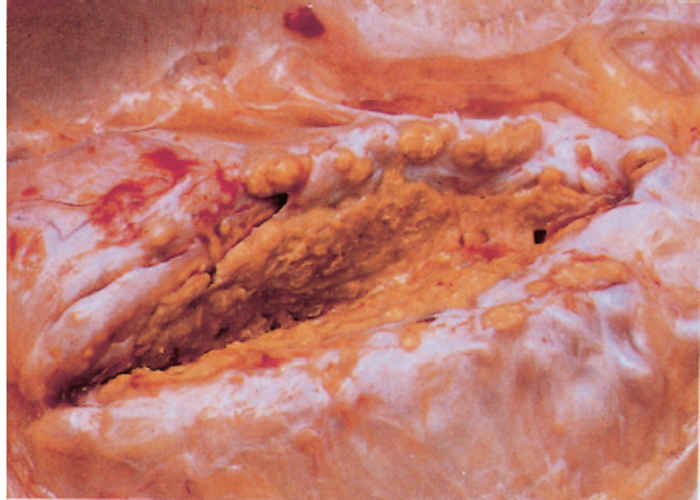
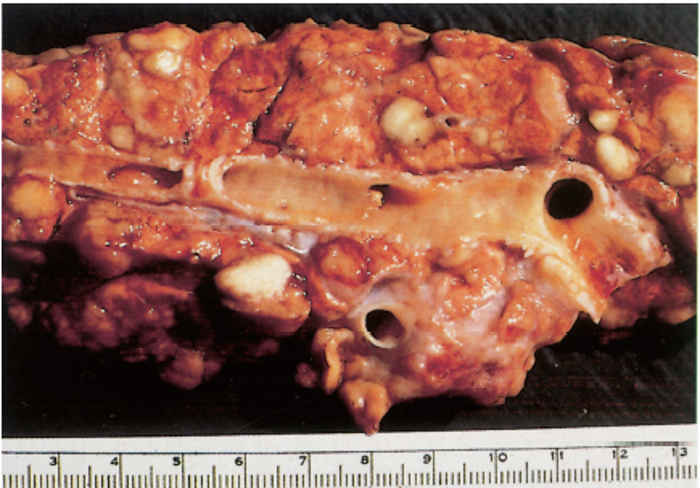
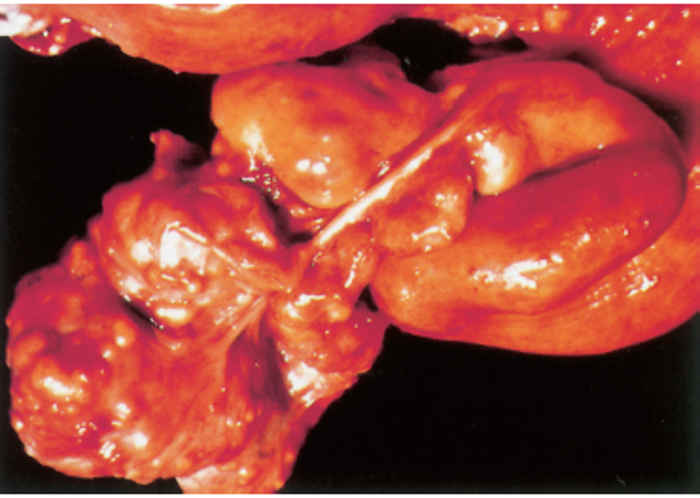
Since most mycobacterial infections are contracted by inhalation or ingestion, tubercles occur predominantly in the retropharyngeal, bronchial, mediastinal and mesenteric lymph nodes (Figures 196.3 to 196.6). The presence of lesions in these sites represents the typical distribution of the lesions encountered in countries in which the disease is actively controlled. In countries where control is either less stringent or not applied, the distribution of lesions may be expected to conform to those typically described for the disease. In adult cattle and older calves, the lungs are affected in 90 to 95 per cent of cases, while in 5 to 10 per cent of cases the digestive tract is infected. In young calves, omphalogenic infection results in hepatic lesions in 49 per cent of cases, while 38 per cent and 13 per cent of cases, respectively, suffer from pulmonary and digestive tract infections. 21 Primary pulmonary lesions can be very small and are easily overlooked. They are located principally subpleurally and caudo-dorsally, but may occur in any lobe and are most successfully detected at necropsy or in the abattoir by palpation of the lungs. Initially, lesions appear macroscopically as partially or completely encapsulated, yellowish foci of caseous necrosis, parts of which may be calcified (Figure 196.7). Encapsulation is not always absolute, which allows local spread through the airways (Figure 196.8). Several such foci may be clustered within a lobule to produce a clover- leaf appearance. Coalescence and expansion of these pneumonic foci result in the development of large areas of a caseating bronchopneumonia (Figure 196.9). In time, the pneumonic focus may become calcified and encapsulated. Bronchial and tracheal ulceration may result in intrapulmonary spread. Pulmonary lesions may heal completely but metastatic lesions in regional lymph nodes that are invariably present, persist.
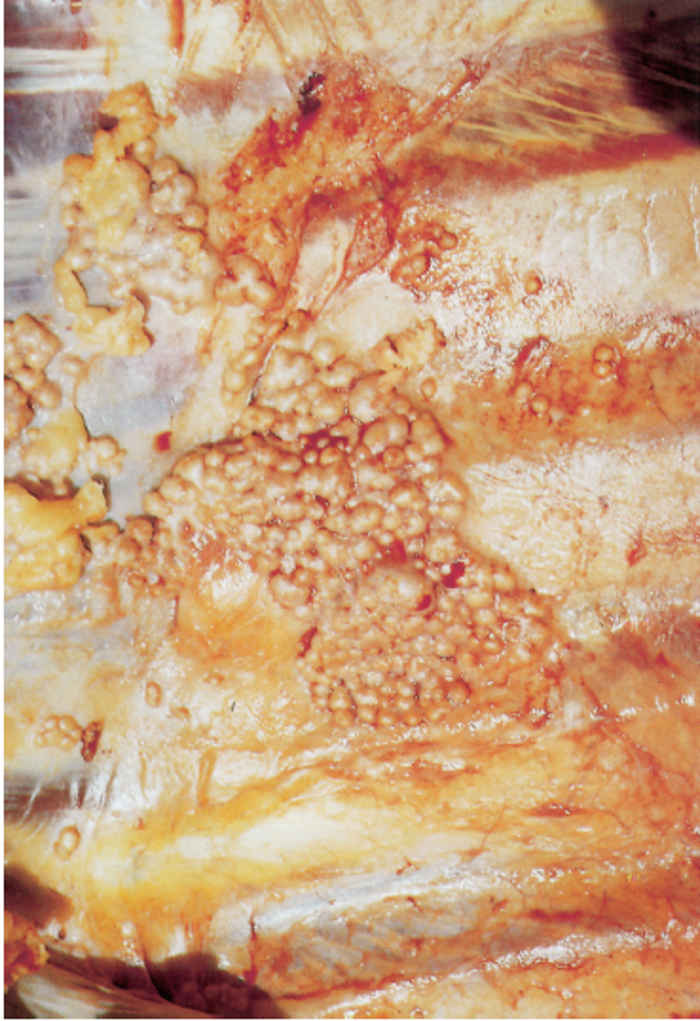
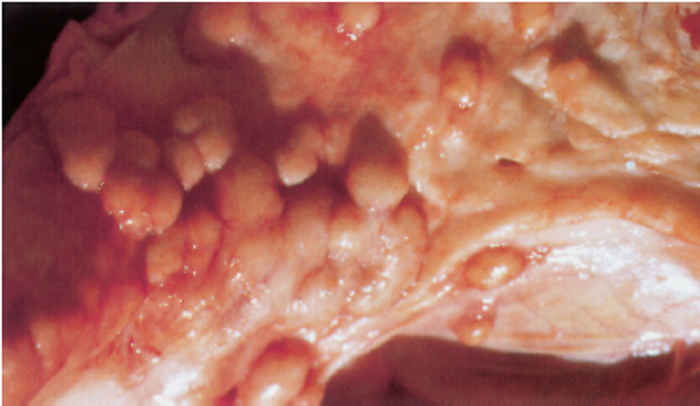
On the pleura or mesentery, spread is aided by respiratory or intestinal peristaltic movements respectively, to cause diffuse, caseous, plaque-like or clustered, protruding, nodular lesions (Figures 196-10 and 196.11). These lesions are referred to as ‘TB-grapes’, and after calcification, as ‘TBpearls’.
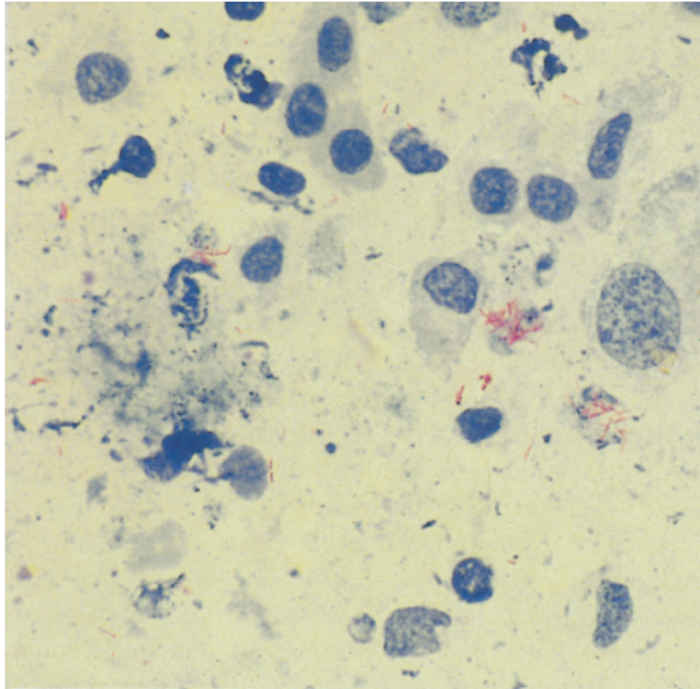
Generalized tuberculosis is characterized by the presence of ‘miliary’ or larger lesions in organs and lymph nodes throughout the body. Miliary lesions are small (from microscopic to 5mmin diameter), and are translucent in the early stage, but become caseous and calcified as they age. In advanced cases of tuberculosis with widespread dissemination, enlarged peripheral lymph nodes, such as the submaxillary, prescapular, precrural and supramammary, may be easily palpable or visible. Single, suppressed, moist coughs may be heard, especially early in the morning, after exercise, or when infected animals are exposed to low environmental temperatures. Dyspnoea may be apparent in advanced cases in which extensive pulmonary lesions have led to impaired respiratory functions135 or where enlarged bronchial lymph nodes cause obstruction of airways. Persistent bloat can occur as a result of pressure on the oesophagus by enlarged mediastinal lymph nodes.12
Hepatic involvement may also follow mycobacteraemia. Lesions may be miliary, although larger, nodular lesions are more common. The kidneys may be similarly involved; miliary lesions are, however, limited to the cortex. Erosion of the renal pelvis and a descending infection may be a sequel.
Tuberculous osteomyelitis, particularly of the vertebrae, ribs and flat bones of the pelvis, occurs in some young animals. The lesions are characterized by extensive caseation and a tendency to liquefy. This exudate may drain through fistulae in the cortex of the affected bone and cause a regional tuberculous myositis. Similarly, articular cartilage may become eroded with the subsequent development of tuberculous arthritis.
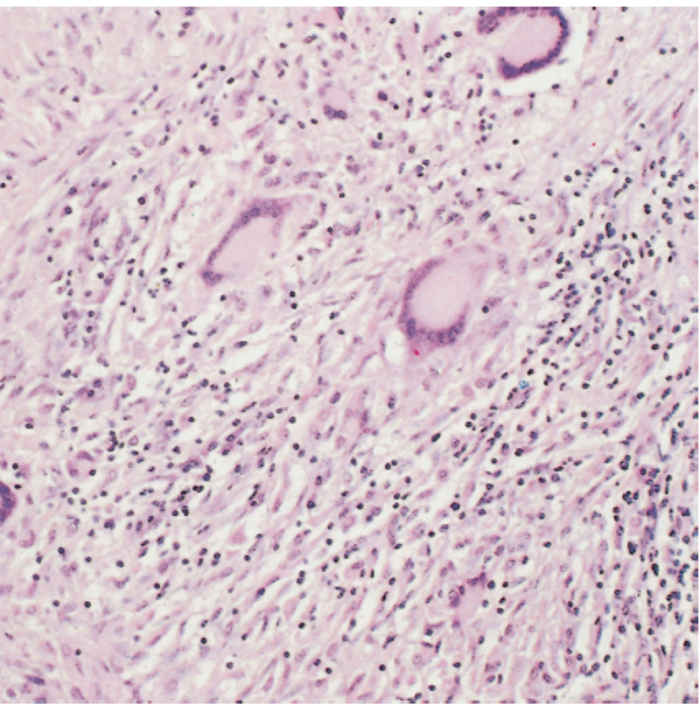
Tuberculous meningitis may follow haematogenous spread or by extension from vertebral lesions. Macroscopically it resembles lesions on serous surfaces, but the lesions are necrotizing and the exudate is often more voluminous.
Lesions in the gastrointestinal tract manifest as nodules or ulcers in the mucosa of the upper alimentary tract, abomasum, or small or large intestine after a per os infection, such as the ingestion of tuberculous milk, or they may be secondary to pulmonary infection following the swallowing of coughed-up, infected material from the lungs. In the intestinal tract, ulcers first develop in Peyer’s patches. The margins of the ulcers are distinct and raised, while the base consists of granulation tissue covered by a dryish, caseous exudate.
Mycobacterial dermatitis is most frequently caused by opportunistic mycobacteria. Single or multiple, often ulcerated, nodules 10 to 80 mm in diameter develop in the skin and subcutis, especially of the legs.156 Upon incision of these nodules, caseous, purulent or possibly mineralized exudate may be exposed. Lesions may involve lymphatics, but the regional lymph nodes are lesion-free — contrary to the situation that pertains in M. bovis infections. Microscopically the lesions consist of typical tuberculous granulomas.
In tuberculous mastitis the affected part of the udder may be firm and/or enlarged with secondary involvement of the supramammary lymph nodes. In some cases, however, enlargement of the lymph nodes in the absence of palpable lesions in the udder itself may indicate the presence of tuberculosis. During the initial stages of infection, the milk appears normal, but in more advanced stages fine floccules may be found in it, which sediment after the milk is left standing in a container, to leave a clear, amber supernatant fluid.11, 143
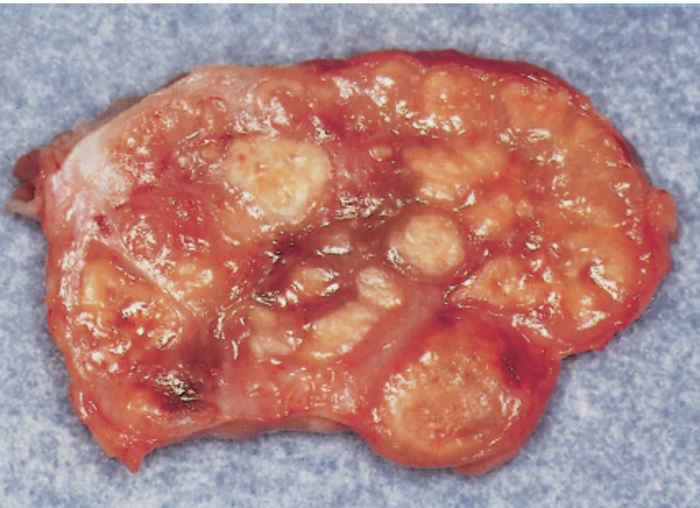
Infection of the bovine udder generally occurs via the haematogenous route. Various forms are encountered, the most common (80 to 90 per cent of cases) being chronic organ tuberculosis,70 in which most of the lobules in an affected gland are replaced by granulation tissue which accentuates the lobulation as the lobules acquire a greyish-red to white colour. Microscopically, lobular outlines are retained but diffuse granulation tissue obliterates the acinar tissue. The intra- and interlobular ducts are always affected, showing marked thickening of the walls due to formation of granulation tissue, while their luminal surfaces tend to caseate. Intramammary spread is solely via the ducts. Contrary to what would be expected, the regional lymph nodes are usually not involved. Disseminated miliary tuberculous mastitis in which nodules of up to 10 mm in diameter are formed, is less common. These lesions occur in clusters and tend to caseate and become calcified. The interlobular ducts are extensively affected. The third form is caseous tuberculous mastitis in which large irregular areas of dry, yellowish, caseous necrotic tissue surrounded by a hyperaemic reaction zone develops. Confluence across interlobular boundaries commonly occurs. Microscopically, there is an exudate composed of fibrin and numerous leukocytes present in the necrotic areas. The fourth manifestation of tuberculous involvement of the mammary gland is tuberculous galactophoritis. In this form, affected lactiferous ducts are dilated and filled with exudate while the acini are relatively uninvolved. The duct walls in these cases are greatly thickened by granulation tissue containing a mononuclear cell infiltration. All cases of mammary tuberculosis are considered to be ‘open’ cases (i.e. excreting mycobacteria in the milk, if the animal is lactating) due to duct involvement.
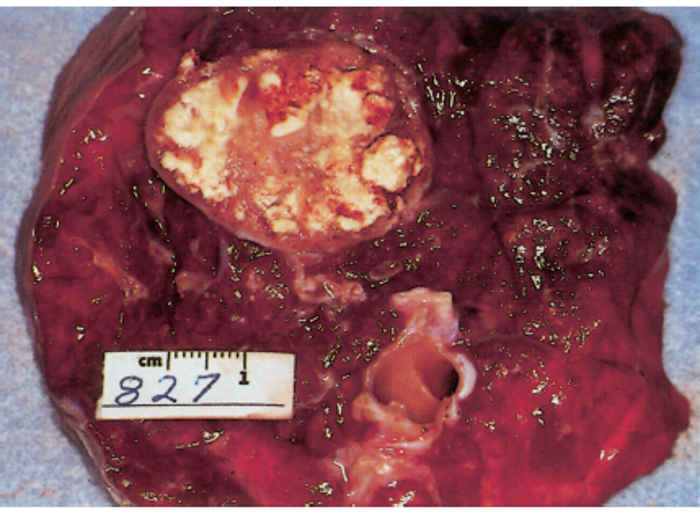
About 1 per cent of new-born calves from tuberculous cows may be born suffering from congenital tuberculosis. Infection of foetuses is generally via the haematogenous route and in most cases secondary to tuberculous endometritis, but it may also be more rarely acquired as the result of either swallowing tuberculous amniotic fluid, which leads to the development of lesions in the intestine or mesenteric lymph nodes, or the inhalation of it, which results in pulmonary disease. Tuberculous lesions in the uterus (Figure 196.12) are usually the result of haematogenous dissemination, but they may originate via the genital route or be an extension of a tuberculous peritonitis. Endometritis as a result of tuberculosis may also cause infertility or result in abortion in the late stages of pregnancy.12 In the majority of congenitally infected calves, the primary complex is found in the liver and portal lymph nodes. As the disease generally progresses quite rapidly and becomes generalized, the young animals die in a few weeks or months. By this time lesions are found particularly in the liver and its regional lymph nodes and in the spleen. As tuberculous lesions are rarely present in the spleen of adult cattle, their presence there is generally regarded as being indicative of congenital infection.
Mycobacterium avium infection in cattle causes a selflimiting disease in which lesions usually do not exceed 20 mm in diameter. Microscopically there is pronounced proliferation of epithelioid cells, formation of giant cells and numerous acid-fast bacilli.90 Mycobacterium tuberculosis infection usually does not progress beyond the development of small granulomas in the pharyngeal, thoracic and mesenteric lymph nodes.
Diagnosis and differential diagnosis
The clinical examination of an animal suspected to be suffering from tuberculosis requires a thorough palpation of all the superficial lymph nodes, the udder in females, and percussion and auscultation of the pulmonary area. A comprehensive investigation of the history of the herd is most important. This should include pertinent questions about the previous introduction of animals into the herd, abattoir reports on animals sent for slaughter, the possibility of bovine tuberculosis on neighbouring properties, and other animal species that may be suffering from bovine tuberculosis. A clinical diagnosis of tuberculosis must be confirmed by the application of the tuberculin test or by laboratory tests. Not all specimens will yield bacteria. In all instances, final confirmation of the diagnosis is dependent on culture and identification of the causative organism.
Tubercle bacilli may be tentatively identified by light microscopy of smears prepared from the content of lesions and stained by the Ziehl-Neelsen method. Final confirmation of the diagnosis is dependent on culturing.
With some exceptions, the biological examination of material suspected to contain mycobacteria, such as suspensions of lesions in tissues, exudates and milk, by the parenteral inoculation of guinea-pigs, which are very susceptible to infection, has become outdated as a diagnostic procedure. Biological examination appears only to be justified in case of urine specimens which have repeatedly been negative on culture, and other body fluids, such as aspirates of pleural exudates, suspected tuberculous material obtained by swabbing (e.g. the larynx, and cerebrospinal fluid which may only contain small numbers of mycobacteria).101 These specimens must, however, also be cultured because of the varying pathogenicity of different species of mycobacteria for laboratory animals. To use guinea pigs as a filter to suppress contaminants in the specimen is unnecessary as other decontamination methods are available today.
Soon after Robert Koch discovered the causative organism of tuberculosis in 1882, it was found that a filtrate of the culture medium in which it had been grown could be used as a diagnostic agent when inoculated into infected animals.122 The reactive substance, later identified as tuberculoprotein and named tuberculin,96, 129 is employed to detect delayed hypersensitivity in animals and humans infected with mycobacteria. Tuberculin itself, when injected parenterally, will not induce hypersensitivity. Tests in England and the Netherlands had shown that PPD (Purified Protein Derivative) tuberculin produced from M. bovis was more specific than PPD tuberculin of M. tuberculosis origin for detecting infections with M. bovis in both cattle and guinea pigs.110, 141, 142 This was confirmed in a field trial in South Africa in 1954.84
As PPD tuberculins consist of a mixture of varying-sized fragments of peptide molecules with different antigenic properties,26, 59, 87, 127 consecutive batches, although produced by the same method, may vary in their potency and specificity. Each newly produced batch is compared to a well-defined standard produced by the same method as the batch under test, and the potency is then expressed in biological units.1 In South Africa, two national or house standards for potency testing are kept at the Onderstepoort Veterinary Institute, namely:
- National standard for PPD of bovine tuberculin is standardized against the European Community (EC) standard for PPD of bovine tuberculin in homologously sensitized guinea pigs and in naturally infected tuberculous cattle.1, 36, 60 Until 1986 the EC standard was the only standard for PPD of bovine tuberculin to which a unitage had been assigned. It contains 50 000 Community Tuberculin Units (CTU)/mg of PPD (EC Council Directive 80/219/EEC).60, 126 Since 1986 an international standard for bovine PPD exists.4, 49, 90 It was tested against the EC standard in an international collaborative assay. It was found to have a relative potency of 65 per cent (32 500 CTU/mg of PPD) of the EC standard. International units are nowadays used to express the potency of bovine PPD.
- National standard for PPD of avian tuberculin is standardized against the international standard for PPD of avian tuberculin in homologously sensitized guinea pigs.
The specificity of a batch under test is assayed by a similar method in heterogenously sensitized guinea pigs. Once the test batches are diluted to the strength used for testing cattle and the quality control tests are satisfactory, an expiry date of one year is given.3 Purified Protein Derivative tuberculin should be kept at 4 to 8 °C and be protected from light.85, 86
The intradermal tuberculin test for cattle is based on a specific reaction following the administration of bovine PPD tuberculin.78 For the single intradermal test, an area of skin on the left side of the neck halfway between the juncture of the head and the neck and the fold in front of the shoulder, and halfway between the top and bottom of the neck, is chosen as the inoculation site. For the comparative intradermal test, another similar site is selected, but this is approximately a hand’s breadth more cranial. The hair of the chosen site(s) is removed by shaving (an area of approximately 40 × 40 mm), the skin thickness measured with a calliper, and the required dose (0,1 ml) of tuberculin is injected intradermally using a McLintock syringe. If a comparative tuberculin test is performed, avian tuberculin is injected at the site nearest to the head and bovine tuberculin at the site nearest to the shoulder. The result of the test is read 72 hours after tuberculin has been injected.
The comparative intradermal test is generally used for herds:
- that are tested for the first time,
- in which the tuberculosis status or the history of the herd is unknown or vague,
- in which non-specific reactions occur, or
- when problems have been experienced with the interpretation of the tests.
The simultaneous use of both avian and bovine tuberculin is thus to obtain a definite result in respect of the presence or absence of tuberculosis caused by M. bovis in a herd. The single intradermal tuberculin test, on the other hand, is mainly used:
- where the negative tuberculosis status is known, or
- where herds have already been tested negatively for tuberculosis caused by M. bovis.140
The skin reaction is interpreted according to whether a single or comparative intradermal test was performed and whether the herd history indicates a non-infected or potentially infected herd. The interpretation of the tests is summarized in Table 196.1). As a general guideline, an increase of 4mm and more in the bovine tuberculin reaction or a difference of 5mm between the avian and bovine tuberculin reaction in previous negative herds or 3mm in established positive herds, are regarded as a positive reaction. In the interpretation of the comparative tuberculin test, the reaction is regarded as negative for bovine tuberculosis if the swelling on the injection site of the avian tuberculin exceeds that of the swelling on the site for the bovine tuberculin.140
In an intradermal tuberculin test, a diagnosis is not simply made by measuring the increase in the thickness of the skinfold. If a reaction is present:
- the appearance of the swelling may be round, flat, circumscribed or diffuse,
- it may be red or blue,
- signs of oozing of oedematous fluid and necrosis may be seen,
- swelling of the lymphatic vessels in the skin surrounding the inoculation site may be evident, and
- systemic changes such as shivering, ruffled hair, and listlessness may be observed.
Table 196.1 Diagnostic criteria that are used to interpret skin reactions following intradermal tuberculin tests
| TEST PERFORMED | INCREASE GREATER | MEDIAN* | INCREASE SMALLER |
|---|---|---|---|
| NEGATIVE HERD Bovine tuberculin alone | Suspect | >6 mm< | Negative |
| Comparative test | Suspect (provided the reaction to bovine tuberculin is 4 mm or more) | >+3 mm< (bovine minus avian tuberculin) | Negative |
| POSITIVE HERD Bovine tuberculin alone | Positive | >4 mm< | Negative or suspect |
| Comparative test | Positive (provided the reaction to bovine tuberculin is 4 mm or more) | >+3 mm< (bovine minus avian tuberculin) | Negative or suspect |
* Median is the measurement by which an increase or decrease in skin thickness is judged
The site where each tested animal was injected must always be palpated because reactions may occur that are not clearly visible. Palpation may reveal:
- consistency of the reaction,
- presence of oedema,
- local increase in skin temperature,
- pain, and
- adhesions between the skin and the subcutaneous tissue.
In addition, the state of regional and other superficial lymph nodes should be assessed by palpation. These, especially the superficial cervical (prescapular) and supramammary lymph nodes, may be swollen.
However, it is important to remember that an increase in skin thickness may occur without any of the above signs being present. In such cases one has to rely mainly on the guidelines provided for the increase in skin thickness as well as the herd history to finally make a diagnosis.
It is important to consider all the variables that might influence the final diagnosis and thus the herd is not immediately classified positive merely on the detection of a single ‘positive’ sign.
The interpretation of the test is done on a herd basis. Depending on the previous history of the herd and the results obtained following the visual inspection of the test site of all the animals before each animal is examined individually, the herd is provisionally classified as free or possibly free from tuberculosis, or as infected according to the criteria given in Table 196.1.
Other factors that must be taken into consideration during the interpretation of the intradermal test include the age of the animal, the possibility of infection with species of mycobacteria other than M. bovis (see below) or with other organisms, and a number of physiological factors. In animals recently infected with M. bovis (six weeks to three months) a reasonably typical and large reaction should develop whilst animals that have been infected for a long time (more than six months), will be less sensitive or even anergic.
Mycobacteria spp. including M. tuberculosis, M. paratuberculosis, M. intracellulare (especially in pigs) and M. avium155 and other species grouped according to Runyon’s classification of Group I to Group IV (see the introduction, Mycobacteria: Table 3) have been incriminated as causes of non-specific reactions. For example, M. kansasii of Group I shows a close relationship to M. tuberculosis, whilst the Battey strains from Group III are closely related to M. avium and cause signs in humans that cannot be clinically distinguished from human tuberculosis caused by M. tuberculosis. Mycobacterium fortuitum from Group IV, and other fastgrowers, may sensitize animals.154 Mycobacterium avium var. paratuberculosis is antigenically related to M. avium and accordingly greater reactions are found to avian than to bovine tuberculin.18 Saprophytic mycobacteria which are usually found in food, water and the gastrointestinal tract of herbivores, do play an unimportant role in sensitizing animals, but organisms such as M. smegmatis may cause sensitivity to the intradermal tuberculin test and sometimes may even cause tuberculous mastitis in cattle.113 With the comparative tuberculin test, the sensitivity is more noticeable at the avian site of injection.
Skin lesions often referred to as ‘acid-fast lymphangitis’ or dermatitis nodosa, and caused by saprophytic mycobacteria, appear in the subcutaneous tissue as hard or soft nodules which seldom ulcerate.10 In South Africa, especially in the Western and Eastern Cape Provinces and KwaZulu-Natal, such skin lesions often complicate the interpretation of the tuberculin test.140
Reactions caused by organisms closely related to mycobacteria, such as Norcardia spp., can cause problems in the differential diagnosis of tuberculosis.95, 140 On culture, N. asteroides can also be confused with mycobacteria. Nonspecific reactions can also be caused by conditions such as actinomycosis, actinobacillosis, brucellosis, corynebacteriosis, infections by Trychophyton spp., fascioliasis, peritonitis, abscesses and lumpy skin disease.140
Desensitization may occur in tuberculous cattle immediately before and after calving, resulting in false negative reactions. These animals return again to a positive status four to six weeks later.12
The skin reaction is interpreted according to whether a single or comparative intradermal test was performed and whether the herd history indicates a non-infected or potentially infected herd. The interpretation of the tests is summarized in (Table 196.1).
Tuberculin test reactions can be assessed with a fair degree of accuracy. Although Kleeberg72 estimated that an accuracy of up to 97 per cent is well within South African and international capabilities, and that the application of the single intradermal or comparative tuberculin test will detect a large percentage of the cattle infected with M. bovis, Francis and his colleagues,50 in reviewing the literature, obtained a figure of 81,1 per cent sensitivity; thus, approximately 20 out of 100 infected cattle tested would be missed and would remain within the herd to spread the infection. The success of interpretation is, however, affected by a number of factors.
In cattle that are infected with M. bovis, but for various reasons show insignificant or no reactions at all to intradermal testing and exhibit no clinical signs, false negative diagnoses may be made. The occurrence of false negative, anergic skin test reactions is more common in chronically infected herds where infection may persist for years. During the acute phase of infection, animals usually produce florid, easily detectable skin test reactions. This means that the first whole herd skin test applied to an infected herd is likely to be the most successful test and may identify most if not all of the infected animals. Should an animal become anergic to a skin test it is likely that such an animal has a high percentage chance of remaining negative to subsequent tests. In animals infected for a long time, the intensity of the reaction to the tuberculin test progressively declines.82 In 1 or 2 per cent of the animals in infected herds sensitivity to the tuberculin test may disappear completely.140 The most common conditions leading to false negative diagnoses are the pre-allergic stage of infection, inactive tuberculosis and advanced tuberculosis anergy.
Accumulation of antibodies in the colostrum, prior to and immediately after calving, could explain why infected cows three weeks pre-partum to three to six weeks post-partum, tend to show a decreased sensitivity to the tuberculin test.82, 140 Periparturient stress, though, may be a more logical explanation for increased anergy in pregnant/early lactating cows. Prolonged chemotherapy of infected animals may lead to destruction of all the mycobacteria, leading to the disappearance of the allergic response.72
Reactions that occur following the administration of bovine tuberculin to cattle free from M. bovis are non-specific. 155 The family Mycobacteriaceae embraces many members and cross-reactions are common. The fact does not seriously affect the interpretation of the tuberculin test since there are well-described methods to differentiate these non-specific reactions from reactions due to infection with M. bovis.
As bovine tuberculosis is progressively eradicated from a country, the non-specific reactor problem becomes greater in that the percentage of reactors showing no gross lesions at slaughter tend to increase.155 As only very small lesions may be associated with pronounced reactions to the tuberculin test, a very thorough post-mortem examination may be required to detect them.
In order to detect all positive cases in a herd, retesting is necessary. This will expose certain reactivated anergic cases as well as animals in the pre-allergic stage. Thorough clinical examination, supported by microscopical, bacteriological and biological tests, may also expose some anergic cases.
The results obtained when the caudal skin fold is used as a test site compare favourably with those obtained using the cervical site. For caudal skin fold testing, double strength bovine PPD tuberculin (twice the strength of PPD used in the cervical test) is used. In problem herds containing suspected anergic animals, and/or thick-skinned cattle (Bos indicus breeds), the simultaneous use of both methods (caudal skin fold and cervical) allows detection of up to 6 per cent more true positives and can bring about quicker and more effective eradication of the disease.38, 44, 89, 92, 131
In the subcutaneous tuberculin test, the animals’ rectal temperatures are taken and recorded prior to the injection of tuberculin. Using a conventional hypodermic syringe and needle, an amount of 4ml of PPD tuberculin is injected subcutaneously into the neck of each animal. Temperature measurements of the animals are then taken at two-hourly intervals. A test result is positive when an animal develops a rise in temperature of 1,0 °C or more. This temperature reaction usually develops six to eight hours after administration of tuberculin and reaches a peak at 12 to 24 hours post-injection. 62 A subcutaneous test was regularly used until the early 1930s,129 when it was replaced by the comparative intradermal test, and is only used nowadays in specific situations where anergic animals in infected herds pose a diagnostic problem.
The Stormont test can be applied in a similar manner to detect anergic animals. It is more sensitive than the intradermal test and it is estimated that only 2 per cent of infected animals are misdiagnosed following its application.71 This test is based on the phenomenon that occurs following an intradermal injection of tuberculin in a desensitized, infected bovine: the skin at the original injection site exhibits an increased sensitivity for tuberculin.71, 83 A seven-day interval is allowed between the initial and the ‘boost’ injection, the latter being administered at the same site as the first. An increase in skin thickness of 5mmor more 24 hours after the second injection is considered a positive reaction, irrespective of the type of reaction observed.83
Despite a long and consistent search for suitable serological assays that detect circulating antibody, none of the methods developed have shown sufficient sensitivity or specificity for routine diagnosis. The specificity is further negatively affected by the common antigens shared with other bacterial genera, such as Nocardia and Corynebacterium. For this reason serological tests remain inadequate for clinical application for the diagnosis of the disease.95 However, in the mid- to late 1980s, a new laboratory test based on detection of cell-mediated immune response was developed in Australia.151 The interferon-gamma (IFN-γ) assay is basically a two-phase test that involves the overnight incubation of whole blood with PPD. The following morning the plasma is removed and tested for the presence of the cytokine IFN-γ using a sensitive and specific enzyme immunoassay system. Initial comparisons of the IFN-γ test and tuberculin test in Australia involving more than 6 000 cattle and water buffalo determined that the IFN-γ test had better sensitivity than the tuberculin test, and that maximum sensitivity could be achieved using both tests in parallel. Additional tests to determine the specificity of the assay suggested that specificity was in the order of 98,1 per cent, which is only slightly less than the tuberculin test, and the IFN-γ was accepted in Australia as an official diagnostic test for bovine tuberculosis in 1991. The infrastructure required and the time limits applicable to the assay make its use impractical in many of the developing countries.
The characteristic high sensitivity of the IFN-γ is suited to situations where a high prevalence of tuberculosis exists or when disease is spreading rapidly and it is desirable to detect as many diseased animals as possible. However, since not all diseased cattle are positive in the IFN-γ and the skin test, maximum sensitivity is achieved by the use of both tests in parallel. The IFN-γ assay measures the animal’s immune response to both avian and bovine tuberculins and hence is equivalent to the comparative cervical test. No second visit is required to read the test and there is no post-injection period where the condition of the animals (e.g. stress or poor nutrition) may interfere with the response. In addition, as the IFN-γ is non-invasive, there is no interference with the immune status of the host, and the assay can be repeated more frequently than the skin test, which requires a minimum of 90 days before retesting can be done. In New Zealand, the IFN-γ assay has been shown to be comparable to the comparative skin test in confirming the status of the single intradermal tuberculin test reactors, reducing the length of movement restriction by more than half.119 Numerous field trials performed around the world (e.g. in the Republic of Ireland, Northern Ireland, the USA, Spain, Italy, Brazil and Romania) have confirmed the results of the work done in Australia and New Zealand. The IFN-γ assay has also proven to be useful for the diagnosis of tuberculosis in Asian and African buffalo, goats and kudu. The technique has been evaluated in the testing of buffalo in the Kruger National Park,112 with the advantage that the animals do not have to be recaptured to obtain a result as is required for the skin test.
Because of the insidious nature of the disease and the multiplicity of the rather non-specific clinical signs it evokes, tuberculosis is difficult to diagnose on clinical grounds. The clinical examination of an animal, especially cattle, is aimed more at excluding various other conditions that could be confused with tuberculosis, than actually at diagnosing tuberculosis.
Clinical conditions in cattle that may cause signs similar to bovine tuberculosis are traumatic reticulitis, contagious bovine pleuropneumonia, chronic pasteurellosis, foreign body or aspiration pneumonia, pharyngeal obstruction, actinobacillosis, corynebacteriosis, and various forms of mastitis.
Snoring in cattle may be caused by enlarged tuberculous pharyngeal lymph nodes, but may also be due to the presence of polyps, inflammatory oedema or granulomatous lesions caused by, for example, allergic conditions, fungal infections, actinomycosis or actinobacillosis in the nasal cavity or pharynx.67
Tuberculous mastitis cannot readily be distinguished from certain other forms of mastitis and its presence is not marked until the late stages of the disease. Fibrosis in the case of tuberculous mastitis is most readily detectable at the base of the gland instead of in the vicinity of the cistern as in most other forms of chronic mastitis, and abnormal milk is most commonly discernible at the end of the milking procedure, rather than in the beginning as in many other forms of mastitis.12
A provisional diagnosis of the disease can be made at necropsy or during inspection of carcasses at abattoirs. It must be kept in mind that more than 50 per cent of infected animals can be missed during routine meat inspection.26 In animals that acquired the infection by inhalation, 70 to 90 per cent of lesions are found in either the lymph nodes of the head or in the thoracic cavity. Lesions, however, may occur in any anatomical region of the body. To optimise the process of inspection, the following should be kept in mind: 66 per cent of cattle have only one lesion of which 86 per cent can be detected should only the mediastinal and medial retropharyngeal lymph nodes and the lungs be examined. If, in addition, the parotid, caudal cervical, superficial inguinal and the mesenteric lymph nodes are inspected, 95 per cent of the lesions will be detected.26 The variation in the appearance of lesions of the disease in different animals also makes the detection of lesions difficult for the uninitiated and for those who do not deal with the disease routinely. In addition, the granulomatous lesions of tuberculosis may be confused with those caused by infectious agents such as fungi, staphylococci, Actinomyces and Actinobacillus spp., or foreign bodies, or with abscesses, particularly those containing inspissated and calcified pus, such as those caused by Actinomyces spp. Generalized cases in which serosal involvement with the development of ‘pearls’ or ‘grapes’ has occurred, may be confused with mesothelioma. In most cases where granulomas or other lesions are observed their cause should be determined by direct examination of smears made from the exudate after appropriate staining, and by histopathological and/or cultural methods. Smears prepared from the exudate of lesions or from the deposit of centrifuged milk samples may reveal the presence of acidfast bacilli after they have been stained by the Ziehl-Neelsen method. Histopathological examination of lesions may be used as a routine method for confirmation of the diagnosis in animals that have died and from which specimens have been collected and preserved in 10 per cent formalin for diagnostic purposes. It should, however, be kept in mind that the diagnosis must be based on the detection of acid-fast bacilli in the sections, and that the species of Mycobacterium cannot be identified in such preparations. In addition, the number of bacteria that can be detected in granulomas tends to vary markedly; in those caused by saprophytic mycobacteria, masses of acid-fast bacteria may be seen, whereas in many instances of infection by M. bovis, very few or even no bacteria may be detected even after prolonged examination of sections. The inability thus to detect acidfast bacteria granulomatous lesions in tissue section prepared from specimens obtained from animals suspected to be suffering, or had suffered, from tuberculosis, does not warrant a negative diagnosis. Polymerase chain reaction techniques may in time prove to be as reliable and a more rapid method of detecting acid-fast bacteria in specimens obtained from infected animals.158
Control
Several countries which today are considered to be ‘developed’ began implementing large-scale campaigns to eradicate tuberculosis in cattle not long after Robert Koch’s discovery of the tubercle bacillus. In many of them the bovine disease has been eradicated and human tuberculosis caused by, M. bovis has diminished greatly.
Modern campaigns to eradicate tuberculosis can be extremely effective but potentially very expensive to complete. The general international criteria applicable to the eradication of and acquiring the status of ‘free of bovine tuberculosis’ are contained in the International Health Code of the OIE (Office International des Epizooties) that is updated on a regular basis and available on their website.6
Earlier campaigns took 30 or 40 years to achieve the results that can be obtained within a decade by the application of modern techniques. Nowadays, a few cases occur, sometimes originating from a single source, and point epidemics originating on a single farm or from a single animal may not be noticed in the absence of an ongoing national surveillance system for bovine tuberculosis.
The success of a campaign will depend largely upon the political will and stability within a country as these affect the availability of adequate funding resources. The success also relies on having experienced veterinarians, other animal health administrators, and reliable and efficient laboratory diagnostic services. It should also be recognized that the occurrence of tuberculosis in wildlife is one of the biggest barriers to the success of eradication. The presence of these sources of infection will necessitate the development of new control strategies whose success will, in all probability, be dependent on the future availability of effective vaccines.
The main purpose of campaigns against bovine tuberculosis is the eradication of the disease from a country. A policy which aims at total eradication, as is the case in the USA, Australia and the UK, would be ideal, but the conditions under which animal husbandry is practised in many regions of the developing world make such a policy impracticable. 129 The two most widely used methods of control are the test and immediate slaughter method, and the Bang method, which comprises the isolation of infected animals and the creation of a ‘clean’ herd by the gradual elimination of cattle from the infected herd.
A total slaughter-out policy is only implemented when the initial infection rate proves to be so high (30 per cent and more) that the cost of further testing will not serve the purpose of eradication of the disease.
Eradication on a national scale can only be achieved if there is:
- full control over all movements of cattle,
- compulsory identification of all cattle,
- payment of an incentive to owners for the slaughter of positive reactors,
- compulsory testing of all cattle within specified intervals,
- establishment and maintenance of disease-free areas, with the eventual aim of incorporating the whole country, and
- sufficient funds and manpower to fulfil the task.
In certain deserving cases, for instance in the case of pedigree animals or when a large number of valuable commercial animals are infected, they may be treated with isonicotinic-hydrazide (INH). The value of INH treatment not only lies in its therapeutic effect (a 78 per cent bacteriological cure is possible), but also in its prophylactic value which is as important in the control of the disease as its curative effect. As all reactors that are treated are not cured bacteriologically, INH treatment cannot be regarded as a means of eradicating the disease and the use of therapeutic antibiotic therapy should not be used to provide false hope in disease management. Therapy with INH alone would be justified in situations where an attempt is being made to salvage animals with unique genotypes.
Treatment with INH is regarded as being merely a temporary measure; postponement of the slaughter of positive reactors being granted for a maximum of five years.
The effectiveness of INH treatment in cattle is superior to that of any other drug. It is cheap, easy to administer and has few adverse effects, and at the dosage rate of 10 mg/kg body weight daily for eight weeks, it is highly effective. Intermittent INH medication is less effective than daily dosing. Isonicotinic-hydrazide treatment has been shown to be successful even in situations in which there is an unfavourable combination of high exposure, high virulence and low host resistance.75
Daily INH treatment for one month is insufficient to rid infected animals of tubercle bacilli, but two to three months’ treatment results in the elimination of viable tubercle bacilli from most cases. Following successful chemotherapy, skin reactions to the administration of tuberculin gradually decrease and cease after five months.74, 79 In one experiment in which tuberculous animals were treated with 25 mg/kg body weight per day of INH for 60 days,88 skin reactions to tuberculin became negative after two to four months post-treatment, and 96,4 per cent of the treated animals were considered cured with no lesions being found in those that were slaughtered.
Pasteurization is recognized as a successful method to curtail the spread of the disease to humans and other animals that are fed cows’ milk. Short-time, high-temperature pasteurization is not always effective in destroying tubercle bacilli, even when the milk is retained in the holder for 20 to 25 seconds, rather than the prescribed minimum of 15 seconds at 72 °C. Only those pasteurization plants with flow diversion valves will destroy all tubercle bacilli in milk. The efficacy of pasteurization also depends on the quality of the milk. When the cell content in milk is high or if it contains pus, is sour or is incompletely clarified, efficacy is lower. Milk products such as sour milk, buttermilk, yoghurt and cream cheese made from inadequately pasteurized milk may contain tubercle bacilli, which can remain viable for 14 days after their preparation. In butter, they may remain viable for as long as 100 days. When modern equipment is used, the pasteurization of milk is an inexpensive procedure, but it should be done in strict conformity with health control regulations.
On infected properties, mechanical, physical and chemical agents are used to render rooms, materials, fluids and other substances non-infectious. The most effective methods for destroying mycobacteria are those that are based on the use of heat, such as hot air, burning, cooking, pasteurization, and running or pressurized steam. Ultraviolet rays are biologically active at 21 to 33 °C and tubercle bacilli are killed within 20 minutes after they have been exposed to direct sunlight. Although the germicidal effect of artificially produced ultraviolet light on mycobacteria is utilized in some circumstances in laboratories and elsewhere, its pronounced toxic effect on the skin and eyes of humans requires that only indirect irradiation be used.
Whatever the means of disinfection, it should be followed by very thorough physical cleaning of ceilings, walls, floors, tools, utensils, and furniture, using as large a quantity of water as possible. Inaccessible areas should be treated with a spray disinfectant. Premises and utensils must be disinfected several times at the beginning of any procedure. Final disinfection is done after the last tuberculin reactor has been removed from the herd. Tubercle bacilli are more resistant to chemical action than many other species of vegetative bacteria.
Surfaces to be disinfected should be in contact with a disinfectant that has been demonstrated to be effective in destroying mycobacteria for at least 10 minutes. In some countries’ regulations 3 per cent formalin and in others 2 per cent lysol or 2,5 per cent phenol are prescribed as the standard disinfectants. Phenol disinfectants are, on the whole, more effective than most substances, but they are difficult to dissolve in water and have a strong odour. Simple phenols are irritating to the skin. In the field, a 1 to3per cent concentration of activated chloramine is recommended. Inorganic acids, alkaline substances, quaternary ammonium compounds and chlorides are ineffective. Cresols are less toxic and have some cleansing effect and good penetration but do not dissolve readily in hard water. Iodophors are potent and non-irritant to the nose and eyes, but they taint the skin, are expensive, and must not be mixed with soaps and alkaline cleaners.
Tuberculosis and mycobacteriosis in pigs
Mycobacterial infection of pigs is caused by M. tuberculosis, M. bovis and M. avium.56 Serovars 1, 2 and 3 of the M. avium complex are the cause of tuberculosis in fowls. Poultry infected with serovars 1 and 2 used to be a source of infection for pigs in Europe, where poultry and pigs were kept together. These two serovars (as well as serovar 3) have, however, seldom been found in poultry in South Africa—hence infection with serovars 1 and 2 is rarely found in pigs in South Africa.
In South Africa, most strains isolated from the lymph nodes of pigs belong to theM. avium complex but are serotypes other than serovars 1, 2 and 3,78 while M. tuberculosis, 47 and particularly M. bovis, may also be associated with porcine tuberculous lymphadenitis.117 Since the mid-1960s a high prevalence rate of mycobacterial lymphadenitis in pigs maintained in intensive husbandry systems has been associated chiefly with organisms of the M. avium complex, predominantly serovars 4 and 8, whereas serovars 4, 8, 9, 10, 13, 14, 16 and 17 have been found in the environment.53, 97, 107 The prevalence of mycobacterial lymphadenitis due to infection with serovars of the M. avium complex can become alarmingly high and may cause severe economic loss. Since the environment is the source of this infection, it is difficult to control, and the problem persists today in many piggeries in South Africa and in countries all over the world.
Epidemiology
Mycobacterial lymphadenitis is a well-known condition in pigs.98 An increased prevalence, which may be more pronounced in summer, is often experienced but is restricted to a few piggeries, presumably due to increased levels of environmental contamination by the bacteria.
The maintenance hosts of the serovars 1, 2 and 3 are poultry and birds, but many healthy, rural persons in South Africa have also been found to harbour serovars of these bacteria in small numbers in their respiratory tracts.107 It has been suggested that mycobacteria in faeces may be a method of spread from animal to animal but in comparison to the large numbers of these mycobacteria in the environment which appears to be the main source,78 faecal spread appears to be of little practical significance. Mycobacterium bovis is contracted by the ingestion of infected products such as meat and milk.
During the five-year period 1987 to 1992, 351 mycobacteria were isolated from 552 porcine lymph nodes in the tuberculosis laboratory of the Onderstepoort Veterinary Institute, South Africa. Mycobacterium bovis accounted for 2,5 per cent, M. avium for 75 per cent, and M. tuberculosis for 22 per cent of these mycobacterial isolates. The porcine specimens from which M. tuberculosis was isolated derived from the Western and Eastern Cape provinces in South Africa, where the prevalence of tuberculosis in certain sections of the human population seems to be extremely high. Mycobacterium tuberculosis infection is due to contact between pigs and tuberculous humans or to ingestion of food contaminated with M. tuberculosis.52 Wild pigs are considered a ‘spill-over host’ for M. bovis in Australia, Europe and New Zealand.108
Pathogenesis, clinical signs and pathology
Infection almost exclusively occurs by ingestion which generally results in the development of an incomplete primary complex in lymph nodes regional to the pharynx or intestine, although lesions do rarely occur in the wall of the intestine, and elsewhere due to haematogenous spread.137 Respiratory and wound infections are uncommon.21
The tuberculous lesions are generally only small foci, 1 to 10 mm in diameter, in a few lymph nodes, and are unlikely to cause detectable clinical signs. Generalization with localization of lesions does occur, the clinical signs being dependent on the localization of the lesions.
Lesions in mucosae are generally microscopic, while those in the regional lymph nodes are often grossly visible. Lesions vary according to the type of mycobacterium involved. Mycobacterium bovis causes caseocalcerous tubercles similar to those in cattle, while M. avium causes proliferative, lardaceous, almost sarcoma-like granulomas due to a diffuse multiplication of epithelioid and giant cells as well as fibroplasia without distinct encapsulation of the affected parts. Acid-fast bacteria are present in these lesions. 99
Pulmonary lesions, when they do occur, are usually of haematogenous origin and miliary in nature. Mycobacterium bovis may cause a consolidated, caseous bronchopneumonia due to a confluence of tubercles, while infection with serovars of the M. avium complex may result in the development of miliary lesions along the course of lymphatics in the lungs.
Hepatic lesions caused by M. bovis infection are either miliary or consist of larger nodules, while those due to M. avium infection resemble the parasitic hepatitis caused by ascarid migration, with no typical tubercles being formed. Splenic lesions consisting of bulging nodules are common in generalized tuberculosis, while involvement of serous membranes is rare. Meningeal lesions are usually nodular. Mycobacterium bovis infection may result in localized lesions in bones. Lesions rarely occur in the genitalia, eyes and skin.
Tuberculous lesions in pigs slaughtered in abattoirs are usually limited to one or more lymph nodes. Lesions occur predominantly in the mesenteric nodes, less frequently in the nodes of the head and rarely in the bronchial nodes. Lesions vary in appearance from small, yellowish-white caseous foci 1 to 10 mm in diameter41 to diffuse enlargement of the entire node. Histologically, M. avium complex infection is usually characterized by a diffuse proliferation of epithelioid and giant cells. Calcification may occur but is generally not as extensive as that produced by mammalian mycobacteria. The same applies to encapsulation of the lesion by connective tissue. The histopathologic differentiation of mycobacterial lymphadenitis caused by mammalian or avian complex organisms is not considered possible.64
Diagnosis
The use of the comparative tuberculin test to some extent does assist in identifying the presence and type of infection. 64 The intradermal tuberculin test in pigs is performed by injecting 0,01 ml of standard potency mammalian tuberculin into the skin fold at the base of the ear. It is, however, relatively inaccurate in this species.12
Lesions in lymph nodes detected macroscopically are not diagnostic, but are usually highly indicative of tuberculous lymphadenitis. Further confirmation of their tuberculous nature is based on their histological appearance and the presence of acid-fast bacilli which may be demonstrated in tissue sections or smears prepared from exudates and stained by the Ziehl-Neelsen method. Final identification of the species of mycobacterium concerned is done by cultural means. This latter procedure can take up to three months to perform and is, therefore, of no immediate diagnostic assistance. It is, however, of value to producers experiencing financial losses as a result of the condemnation at abattoirs of organs or body parts, such as heads, in identifying the source of the infection, particularly if the responsible organism should prove to be M. tuberculosis or M. bovis.
Sensitized lymphocytes and mycobacterial antibodies detectable by application of ELISA have been reported to occur 14 to 28 days after exposure to serovars of the M. avium complex or to M. bovis.104, 133
Differential diagnosis
Tuberculosis-like lesions in the lymph nodes of pigs may be confused with lesions caused by Rhodococcus equi20 and with streptococcal lymphadenitis (jowl abscess) caused by Streptococcus porcinus which is characterized by abscessation of submaxillary lymph nodes and occasionally deeper nodes as a result of haematogenous spread.
Control
The routine control of infection lies in protecting pigs from contact with tuberculous humans or products originating from infected cattle, such as dairy products. The feeding of uncooked swill originating from mine compounds, army camps and especially hospitals poses obvious dangers. When cultural evidence shows the infection to be of environmental origin, as is usually the case on most high-prevalence farms, the difficulties inherent in controlling or eradicating M. avium complex infection are formidable indeed. These opportunist organisms appear to enjoy an ubiquitous and free-living existence in nature, and are resistant to antituberculous drugs. Moreover, they may occur in profusion on high-prevalence premises, particularly in zones with a temperate or subtropical climate.98 With a yield of 18 to 250 mycobacterial colonies per feed sample, it has been deduced that a pig eating 3 kg of such feed per day would ingest 5 000 to 75 000 of these mycobacteria. It is therefore not surprising that some investigators have found these organisms in the faeces of pigs.78 Wood shavings and sawdust have been found to be heavily contaminated with M. avium complex organisms in South Africa and other countries, including the UK,78, 150 so that one proven method of reducing or even eliminating such infection is by dispensing with bedding that consists of the by-products of the timber industry.
It is not known whether BCG vaccination has any protective capacity against M. avium-intracellulare infection in pigs.35
Tuberculosis in sheep and goats
Tuberculosis is less common in goats than in cattle and is rare in sheep, and generally occurs when these species are exposed to infected cattle. However, it has recently been reported to occur in milking goat flocks in Spain, where mortality rates varied from 20 to 50 per cent.51 Tuberculosis with a flock prevalence of more than 30 per cent has been diagnosed in feral goats in New Zealand.120 In Spain, goats appear to be a maintenance host for tuberculosis and most cases are caused by a variant of M. bovis recently classified as M. tuberculosis subsp. caprae.7, 58
The lesions of tuberculosis in goats and sheep relatively closely resemble those in cattle. Infection mostly occurs via the respiratory route and lesions in almost all the cases occur in the thoracic cavity and lungs.21
In sheep, it is only possible to do satisfactory tuberculin tests on skin depilitated with barium sulphide paste, while in goats, satisfactory tests (such as the standard intradermal tuberculin test) can be performed if the hair is clipped short.116
Tuberculosis in horses
Tuberculosis is rare in horses, and infections that do occur are mostly due to M. bovis. Serotypes of the M. avium complex and M. tuberculosis, however, may also infect horses.46 The route of infection is almost solely alimentary, resulting in lesions often only in the retropharyngeal and/or mesenteric lymph nodes.
Fatal cases are usually due to a generalized miliary or multifocal infection involving the lungs, liver, spleen, and serous membranes. Allegedly, lesions occur commonly in cervical vertebrae but no proof exists to substantiate this.39 In this form, painful osteomyelitis develops which results in a stiffness of the neck and an inability to eat off the ground.12 Less common signs include pyuria, coughing due to pulmonary lesions, lymph node enlargement, nasal discharge and a fluctuating temperature.
Tubercles in horses are uniform, smooth, greyish, lardaceous, sarcoma-like granulomas that rarely undergo caseation and calcification. Microscopically, they consist of epithelioid cells, macrophages, giant cells and lymphocytes, and become more fibrous as they age. Organisms are scarce in these lesions.
Pulmonary infection is usually miliary and the lesions manifest as firm, ‘glassy’ nodules. The bronchial lymph nodes are enlarged and have a sarcoma-like appearance. In intestinal tract infections ulcers occur in the large intestine in particular. Mycobacterium avium may produce a proliferative enteritis not unlike that of Johne’s disease in cattle. Lesions in the liver, spleen and serosae are large lardaceous nodules.13 Serosal involvement causes a copious effusion.
Horses appear to be much more sensitive than cattle to tuberculin, and probably much smaller doses of standardized tuberculin are required when conducting an intradermal test with it than is the case in cattle. In horses, the results obtained with subcutaneous and intradermal tuberculin tests are very erratic and must be assessed with caution, especially when the test is positive.12
References
- ANON., 1968. Expert Committee on Biological Standardization. Twentieth Report. World Health Organization Technical Report Series, 384, 23–41. Geneva: WHO.
- ANON., 1977–1988. Annual Reports of the Director of Veterinary Services, Namibia.
- ANON., 1984. Manual for the Production of PPD of Bovine and Avian Tuberculin. Section of Diagnostic Antigens, Onderstepoort Veterinary Institute, Pretoria, South Africa.
- ANON., 1987. Expert Committee on Biological Standardization. Thirty-sixth Report. World Health Organization Technical Report Series, 745. World Health Organization, Geneva 1987.
- ANON., 2001. World Animal Health in 2001. Volumes 1 & 2. Office International des Epizootics (OIE).
- ANON., 2002. OIE Website: www.oie.int/eng/normes/en—norm.htm
- ARANAZ, A., LIÉBANA, E., GOMEZ-MOMPASO, E., GALAN, J.C., COUSINS, D., ORTEGA, A., BLASQUEZ, J., BAQUERO, F., MATEOS, A., SUAREZ, G. & DOMINGUEZ, L., 1999. Mycobacterium tuberculosis subsp. caprae sp. nov.: A taxonomic study of a new member of the Mycobacterium tuberculosis complex isolated from goats in Spain. International Journal of Systematic Bacteriology, 49, 1263–1273.
- ARTOIS, M., DELAHAY, R., GUBERTI, V. & CHEESMAN, C., 2001. Control of infectious diseases of wildlife in Europe. Veterinary Journal, 162, 141–157.
- BENGIS, R., KRIEK, N.P.J. & HUCHZERMEYER, H.F.A.K., 1991. State Veterinarian, Skukuza, South Africa. Unpublished data.
- BENJAMIN, R.G. & DANIEL, T.M., 1982. Serodiagnosis of tuberculosis using the enzyme-linked immunosorbent assay (ELISA) of antibody to Mycobacterium tuberculosis antigen 5. American Review of Respiratory Diseases, 126, 1013–1016.
- BERNARDELLI, A., BASTIDA, R., LOUREIRO, J., MICHELIS, H., ROMANO, M.I., CATALDI, A. & COSTA, E., 1996. Tuberculosis in sea lions and fur seals from the south-western Atlantic coast. Revue Scientifique et Technique – Office International des Epizooties, 15, 985–1005.
- BLOOD, D.C., RADOSTITS, O.M. & HENDERSON, J.A., 1989. Diseases caused by bacteria. IV. In: Veterinary Medicine. 7th edn. London: Baillière, Tindall & Cox.
- BUERGELT, C.D., GREEN, S.L., MAYHEW, I.G., WILSON, J.H. & MERRITT, A.M., 1988. Avian mycobacteriosis in three horses. Cornell Veterinarian, 78, 365–380.
- CARMICHAEL, L., 1941. Bovine tuberculosis in the tropics with special reference to Uganda. Part II. Experimental infection in zebu cattle. Veterinary Journal, 97, 329–339.
- CARTER, C., CREWS, K., HOYLE, F. & RYAN, T., 1995. The movement of cattle from herds in which Mycobacterium bovis has been diagnosed. In: griffin, f. & de lisle, g., (eds). Tuberculosis in Wildlife and Domestic Animals. Conference Series No. 3, Dunedin, New Zealand. Otago: University of Otago Press. pp. 298–299.
- CASSIDY, J.P., BRYSON, M.M., GUTIÉRREZ, C., FORSTER, F., POLLOCK, J.M. & NEILL, S.D., 2001. Lymphocyte subtypes in experimentally induced early-stage bovine tuberculosis lesions. Journal of Comparative Pathology, 124, 46–51.
- CHEESEMAN, C.L., WILESMITH, J.W. & STUART, F.A., 1989. Tuberculosis: The disease and its epidemiology in the badger, a review. Epidemiology, 103, 113–125.
- CHIODINI, R.J., VAN KRUININGEN, H.J. & MERKAL, R.S., 1984. Ruminant paratuberculosis (Johne’s disease): The current status and future prospects. Cornell Veterinarian, 74, 218–262.
- CLANCY, J.K., 1977. The incidence of tuberculosis in lechwe (Marsh antelope). Tubercle, 58, 151–156.
- CLAPP, H.K., 1956. Tuberculosis-like lesions in swine in South Australia. Australian Veterinary Journal, 32, 110–113.
- COHRS, P., 1967. Textbook of the Special Pathological Anatomy of Domestic Animals. Oxford: Pergamon Press.
- COLLINS, C.H. & GRANGE, J.M., 1983. A review: The bovine tubercle bacillus. Journal of Applied Bacteriology, 55, 13–29.
- COLLINS, C.H. & GRANGE, J.M., 1987. Zoonotic implication of Mycobacterium bovis infection. Irish Veterinary Journal, 41, 363–366.
- COLLINS, C.H., GRANGE, J.M. & YATES, M.D., 1985. Organization and Practice in Tuberculosis Bacteriology. London, Boston, Durban, Singapore, Sydney, Toronto, Wellington: Butterworths.
- COOKE, M.M., GEAR, A.J., NAIDOO, A. & COLLINS, D.M. 2002. Accidental Mycobacterium bovis infection in a veterinarian. New Zealand Veterinary Journal, 50, 36–38.
- CORNER, L.A., 1994. Post mortem diagnosis of Mycobacterium bovis in cattle. Veterinary Microbiology, 40, 53–64.
- COSIVI, O., GRANGE, J.M., DABORN, C.J., RAVAGLIONE, M.C., FUJIKURA, T., COUSINS, D., ROBINSON, R.A., HUCHZERMEYER, H.F.A.K., DE KANTOR, I. & MESLIN, F.-X., 1998. Zoonotic tuberculosis due to Mycobacterium bovis in developing countries. Emerging Infectious Diseases, 4, 59–70.
- COUSINS, D.V. & DAWSON, D.J., 1999. Tuberculosis due to Mycobacterium bovis in the Australian population: Cases recorded during 1970–1994. International Journal of Tuberculosis and Lung Diseases, 3, 715–721.
- COUSINS, D.V., FRANCIS, B.R., GOW, B.L., COLLINS, D.M., MCGLASHAN, C.H., GREGORY, A. & MACKENZIE, R.M., 1990. Tuberculosis in captive seals: Bacteriological studies on an isolate belonging to the Mycobacterium tuberculosis complex. Research in Veterinary Science, 48, 196–200.
- COUSINS, D.V., WILLIAMS, S.N., REUTER, R., FORSHAW, D., CHADWICK, B., COUGHRAN, D., COLLINS, P. & GALES, N., 1993. Tuberculosis in wild seals and characterisation of the seal bacillus. Australian Veterinary Journal, 70, 92–97.
- DANIEL, T.M. & JANICKI, B.W., 1978. Mycobacterial antigens: A review of their isolation, chemistry and immunological properties. Microbiological Reviews, 42, 84–113.
- DANNENBERG, A.M. JR., 1989. Immune mechanisms in the pathogenesis of pulmonary tuberculosis. Reviews of Infectious Diseases. II. Supplement 2, S369–378.
- DE LISLE, G.W., BENGIS, R.G., SCHMIDT, S.M. & O’BRIEN, D.J.O., 2002. Tuberculosis in free-ranging wildlife: detection, diagnosis and management. Revue Scientifique et Technique—Office International des Epizooties, 21, 317–334.
- DE LISLE, G.W. & HAVILL, P.F., 1985. Mycobacteria isolated from deer in New Zealand from 1970–1983. New Zealand Veterinary Journal, 33, 138–140.
- DEY, B.P., 1986. Mycobacterioses in swine and their significance to public health. USDA. Bibliographies and Literature of Agriculture, 49, 47.
- DOBBELAER, R., O’REILLY, L.M., GENICOT, A. & HAAGSMA, J., 1983. The potency of bovine PPD tuberculin in guinea-pigs and tuberculous cattle. Journal of Biological Standardization, 11, 213–220.
- DUBE, J.G., 1989. Director of Veterinary Services, P.O. Box 162, Mbabane, Swaziland. Personal communication.
- DUFFIELD, B.J., NORTON, J.H. & STREETEN, T.A., 1985. Application of the comparative cervical test to the identification of false positive reactions to the bovine tuberculin caudal fold test. Australian Veterinary Journal, 62, 424–426.
- DUNGWORTH, D.L., 1985. The respiratory system. In: jubb, k.v.f., kennedy, p.c. & palmer, n., (eds). Pathology of Domestic Animals. 3rd edn. Orlando: Academic Press.
- EKDAHL, M.O., SMITH, B.L. & MONEY, D.F., 1970. Tuberculosis in some wild and feral animals in New Zealand. New Zealand Veterinary Journal, 18, 44–45.
- EL MOSSALAMI, E., SIAM, M.A. & EL SERGANY, I.M., 1971. Studies on tuberculosis-like lesions in slaughtered camels. Zentralblatt für Veterinärmedizin, Reihe B, 18, 253–261.
- ELSAID, H.M., 1992. Diagnosis of tuberculosis in buffaloes: Correlation of in vivo and in vitro tests of cell-mediated immunity. Proceedings of the International Conference on Animal Tuberculosis in Africa and the Middle East, Cairo.
- FANNING, A. & EDWARDS, S., 1991. Mycobacterium bovis infection in human beings in contact with elk (Cervus elaphus) in Alberta, Canada. The Lancet, 338, 1253–1255.
- FAUL, A., COLLY, P. & HUCHZERMEYER, H.F.K.A., 1988. Directorate of Animal Health, Private Bag X138, Pretoria 0001. Unpublished data.
- FLAMAND, J., 1993, Natal Parks Board, Mtubatuba, South Africa. Personal communication.
- FLORES, J.M., SANCHEZ, J. & CASTANO, M., 1991. Avian tuberculosis dermatitis in a young horse. The Veterinary Record, 128, 407–408.
- FOURIE, P.J.J., DE WET, G.J. & VAN DRIMMELEN, G.C., 1950. Tuberculosis in pigs caused by Mycobacterium tuberculosis var. hominis. Journal of the South African Veterinary Medical Association, 21, 70.
- FRANCIS, J., 1958. Tuberculosis in Animals and Man. A Study in Comparative Pathology. London: Cassel.
- FRANCIS, J., 1979. Specificity and potency of PPD tuberculins. Discussion Paper for Committee on Tuberculosis in Animals, International Union against Tuberculosis, Paris. October 1979.
- FRANCIS, J., SEILER, R.J., WILKIE, I.W., O’BOYLE, D., LUMSDEN, M.J. & FROST, A.J., 1978. The sensitivity and specificity of various tuberculin tests using bovine PPD and other tuberculins. The Veterinary Record, 103, 420–435.
- GARCIA-MARIN, F., 1993. Report of meetings of IUATLD scientific committee on tuberculosis in animals, Paris. 15–16 October 1992. International Union Against Tuberculosis and Lung Diseases Newsletter. March 1993:14–16.
- GRANGE, J.M. & YATES, M.D., 1994. Zoonotic aspects of Mycobacterium bovis infection. Veterinary Microbiology, 40, 137–151.
- GREATHEAD, M.M., 1969. Porcine mycobacterial lymphadenitis: Meat hygiene considerations. Journal of the South African Veterinary Medical Association, 40, 259–262.
- GRIFFIN, F.J.T., 1988. The aetiology of tuberculosis and mycobacterial disease in farmed deer. Irish Veterinary Journal, 42, 23–26.
- GRIFFIN, J.F.T., MACKINTOSH, C.G. & BUCHAN, G.S., 1995. Animal models of protective immunity in tuberculosis to evaluate candidate vaccines. Trends in Microbiology, 3, 418–424.
- GRIFFITH, F. & GRIFFITH, A.S., 1911. Final Report of the Royal Commission on Tuberculosis, Part 2, Appendix 3, 152. London: HMSO.
- GUILBRIDE, P.D.L., ROLLINGSON, D.H.L., MCANULTY, E.G., ALLEY, J.G. & WELLS, E.A., 1963. Tuberculosis in the free-living African (Cape) buffalo (Syncerus caffer caffer, Sparrman). Journal of Comparative Pathology, 73, 337–348.
- GUTIÉRREZ, M., SAMPER, S., GAVIGAN, J.A., GARCIA-MARIN, J.F. & MARTIN, C., 1995. Differentiation by molecular typing of Mycobacterium bovis strains causing tuberculosis in cattle and goats. Journal of Clincial Microbiology, 33, 2953–2956.
- HAAGSMA, J., 1980. Standardization of bovine tuberculins. Report for the International Union against Tuberculosis, Prague.
- HAAGSMA, J., O’REILLY, L.M., DOBBELAER, R. & MURPHY, T.H., 1982. A comparison of the relative potencies of various bovine PPD tuberculins in naturally infected tuberculous cattle. Journal of Biological Standardization, 10, 273–284.
- HARGREAVES, S.K., 1993. Director of Veterinary Services, P.O. Box 8012, Causeway, Harare, Zimbabwe. Personal communication.
- HENNING, M.W., 1956. Animal Diseases in South Africa. 3rd edn. South Africa: Central News Agency.
- HEWSON, P.I. & SIMPSON, W.J., 1987. Tuberculosis infection in cattle and badgers in an area of Staffordshire. The Veterinary Record, 120, 252–256.
- HIMES, E.M., MILLER, L.D. & THOEN, C., 1983. Swine tuberculosis: Histologic similarities of lesions from which Mycobacterium tuberculosis, M. avium complex or M. bovis was identified. Proceedings of the Twenty-sixth Meeting of the American Association of Veterinary Laboratory Diagnosticians.
- HOFFNER, S.E., BOLSKE, G., HOFFMANN, E. & KÄLLENIUS, G., 1992. Improved detection of Mycobacterium bovis in deer. Proceedings of the International Conference on Animal Tuberculosis in Africa and the Middle East, Cairo.
- HUCHZERMEYER, H.F.A.K., 1992. Onderstepoort Veterinary Institute, Pretoria, South Africa. Unpublished data.
- HUNGERFORD, T.G., 1967. Diseases of Livestock. 6th edn. Sydney: Angus and Robertson.
- HUTCHEON, D., 1880. Tering, consumption, tables mesenterica. Annual Report, Colonial Veterinary Surgeon, Cape of Good Hope.
- JENSEN, K.A., 1953. Bovine tuberculosis in man and cattle. In: Advances in the Control of Zoonoses. WHO/FAO Seminar on Zoonoses. Vienna, November 1952, 11, Geneva: World Health Organization.
- JUBB, K.V.F., KENNEDY, P.C. & PALMER, N., 1985. The female genital system. In: Pathology of Domestic Animals. 3rd edn. Orlando: Academic Press.
- KERR, W.R., LAMONT, H.G. & MCGIRR, J.L., 1946. Further studies on tuberculin sensitivity in the bovine. The Veterinary Record, 61, 466–475.
- KLEEBERG, H.H., 1960. The tuberculin test in cattle. Journal of the South African Veterinary Medical Association, 31, 213–225.
- KLEEBERG, H., 1961. The tuberculin test in cattle. Journal of the South African Veterinary Medical Association, 32, 482–486.
- KLEEBERG, H.H., 1963. Eradication of bovine tuberculosis. Journal of the South African Veterinary Medical Association, 34, 393–400.
- KLEEBERG, H.H., 1967. The use of chemotherapeutic agents in animal tuberculosis. Veterinarian (London), 4, 197–211.
- KLEEBERG, H.H., 1975. Tuberculosis and other mycobacterioses. In: hubbert, w.t., mcculloch, w.f. & schnurrenberger, d.r., (eds). 6th edn. Diseases Transmitted from Animals to Man. Springfield, Illinois: Charles C. Thomas.
- KLEEBERG, H.H., 1981. Epidemiology of mycobacteria other than tubercle bacilli in South Africa. Reviews of Infectious Diseases, 3, 1008–1012.
- KLEEBERG, H.H. & NEL, E.E., 1969. Porcine mycobacterial lymphadenitis. Journal of the South African Veterinary Medical Association, 40, 233–250.
- KLEEBERG, H.H. & WORTHINGTON, R.W., 1963. A modern approach to the control of bovine tuberculosis. Journal of the South African Veterinary Medical Association, 34, 383–391 and 565–568.
- KOLLIAS, G.V., 1978. Clinical and pathologic features of mycobacterial infection in sika and fallow deer. In: montali, r.j., (ed.). Proceedings of a Symposium on Mycobacterial Infections in Zoo Animals. Washington, DC.: Smithsonian Institute Press.
- KRAUSS, H., ROETTCHER, D., WEISS, R., DANNER, K. & HÜBSCHLE, O.J.B., 1984. Wildtiere als Infektionspotential für Nutztiere: Untersuchungen an Wild in Zambia. Giessener Beiträge zur Entwicklungsforschung, Reihe 1, Band 10, Wissenschaftlisches Zentrum Tropeninstitut Justus-Liebig- Universität, Giessen.
- LAMPRECHTS, M.C., 1956. Aspects of tuberculin testing in cattle. Journal of the South African Veterinary Medical Association, 27, 225–243.
- LAMPRECHTS, M.C., 1957. Some observations on the Stormont test. Journal of the South African Veterinary Medical Association, 28, 169–174.
- LAMPRECHTS, M.C., 1958. Report to the Director of Veterinary Services, Onderstepoort Veterinary Institute.
- LANDI, S. & HELD, H.R., 1975. Effect of light on tuberculin purified protein derivative solutions. American Review of Respiratory Diseases, 111, 52–61.
- LANDI, S., HELD, H.R. & GUPTA, K.C., 1975. The multi-facets of tuberculin standardization. WHO: International Abstracts of Biological Sciences Symposium on Standardization and Control of Allergens Administered to Man, Geneva, 1974, Developments in Biological Standardization, 29, 393–411.
- LANDI, S., HELD, H.R. & TSENG, M.C., 1970. Relationship between the molecular weight of tuberculoprotein and its biological potency. Progress in Immunobiological Standardization, 4, 512–520.
- LANGENEGGER, J., CAVALCANTE, M.I. & LIRA, A.D., 1991. Massal treatment of bovine tuberculosis with isoniazid. Pesquisa Veterinária Brasileira, 11, 21–23.
- LARSON, A.B., GROTH, A.H. & JOHNSON, H.W., 1950. Allergic response to johnin and tuberculin of various skin regions of cattle. American Journal of Veterinary Research, 11, 301–303.
- LEE, M.T., 1987. Central Veterinary Laboratory, Weybridge, England. Personal communication.
- LEPPER, A.W.D. & CORNER, L.A., 1983. Naturally occurring mycobacteriosis of animals. In: ratledge, c. & stanford, j., (eds). The Biology of Mycobacteria. Vol. II. London, New York: Academic Press.
- LEPPER, A.W.D., NEWTON-TABRETT, D.A., CORNER, L.H., CARPENTER, M.T., SCANLON, W.A., WILLIAMSON, O.J. & HELWIG, D.M., 1977. The use of bovine PPD tuberculin in the single caudal fold test to detect tuberculosis in beef cattle. Australian Veterinary Journal, 53, 208–213.
- LEPPER, A.W.D. & PEARSON, C.W., 1973. The route of infection in tuberculosis of beef cattle. Australian Veterinary Journal, 49, 266–267.
- LESLIE, I.W. & BIRN, K.J., 1967. Tuberculosis in cattle caused by the avian type tubercle bacillus. The Veterinary Record, 80, 559–564.
- LIND, A. & RIDELL, M., 1984. Immunologically based diagnostic tests: Humoral antibody methods. In: KUBICA, G.P. & WAYNE, L.G., (eds). The Mycobacteria: A Sourcebook. Part B. New York, Basel: Marcel Dekker, Inc.
- LONG, E.R. & SEIBERT, F.B., 1926. The chemical composition of the active principle of tuberculin: I. A non-protein medium suitable for the production of tuberculin in large quantity. American Review of Tuberculosis, 13, 393–397.
- LOVEDAY, R.K., 1968. Swine health under intensification. Journal of the South African Veterinary Medical Association, 39, 117–118.
- LOVEDAY, R.K., 1969. Porcine mycobacterial lymphadenitis: Some epidemiological aspects. Journal of the South African Veterinary Medical Association, 40, 253–256.
- LUKE, D., 1958. Tuberculosis in the horse, pig, sheep and goat. The Veterinary Record, 70, 529–536.
- MAHMOD, K.H., ROOK, G.A.W., STANFORD, J.L., STUART, F.A. & PRITCHARD, D.G., 1987. The immunological consequences of challenge with bovine tubercle bacilli in badgers (Meles meles). Epidemiology and Infection, 98, 155–163.
- MARKS, J., 1972. Ending the routine guinea-pig test. Tubercle, 53, 31–34.
- MEISSNER, G. & SCHRÖDER, K.H., 1974. Bovine tuberculosis in man and its correlation with bovine cattle tuberculosis. Bulletin of the International Union Against Tuberculosis, 49, 145–148.
- MONTALI, R.J., 1978. Proceedings of a Symposium on Mycobacterial Infections in Zoo Animals. Washington, DC.: Smithsonian Institute Press.
- MUSCOPLAT, C.C., THOEN, C.O., CHEN, A.W., RAKICH, P.M. & JOHNSON, D.W., 1975. Development of specific lymphocyte immunostimulation and tuberculin skin reactivity in swine infected with Mycobacterium bovis and Mycobacterium avium. American Journal of Veterinary Research, 36, 1167–1171.
- MYERS, J.A. & STEELE, J.H., 1969. Bovine tuberculosis. Control in man and animals. St. Louis, Missouri, USA.: Warren H. Green, Inc.
- NEILL, S.D., BRYSON, D.G. & POLLOCK, J.M., 2001. Pathogenesis of tuberculosis in cattle. Tuberculosis, 81, 79–86.
- NEL, E.E., 1981. Mycobacterium avium-intracellulare complex serovars isolated in South Africa from humans, swine and the environment. Reviews of Infectious Diseases, 3, 1013.
- O’REILLY, L.M. & DABORN, C.J., 1995. The epidemiology of Mycobacterium bovis infections in animals and man: A review. Tubercle & Lung Disease, 76 Suppl., 1, 1–46.
- PAINE, R. & MARTINAGLIA, G., 1929. Tuberculosis in wild buck living under natural conditions. The Journal of Comparative Pathology and Therapeutics, XL11(1).
- PATERSON, A.B., 1948. The production of bovine tuberculo-protein. The Journal of Comparative Pathology and Therapeutics, 58, 302–313.
- POLLOCK, J.M., MCNAIR, J., WELSH, M.D., GIRVIN, R.M., KENNEDY, H.E., MACKIE, D.P. & NEILL, S.D., 2001. Immune responses in bovine tuberculosis. Tuberculosis, 81, 103–107.
- RAATH, J.P., BENGIS, R.G., BUSH, M., HUCHZERMEYER, H., KEET, D.F., KERNES, D.J., KRIEK, N.P.J. & MICHEL, A., 1995. Diagnosis of tuberculosis due to Mycobacterium bovis in the African buffalo (Syncerus caffer) in the Kruger National Park. In: griffin, f. & de lisle, g., (eds). Second International Conference on Mycobacterium bovis. Tuberculosis in Wildlife and Domestic Animals. Conference Series No. 3, Dunedin, New Zealand. Otago: University of Otago Press. pp. 313–315.
- RICHARDSON, A., 1970. Bovine mastitis associated with Mycobacterium smegmatis and an untypable mycobacterium. The Veterinary Record, 86, 497–498.
- RIDLEY, D.S., 1983. The histopathological spectrum of the mycobacterioses. In: Biology of Mycobacteria. Vol. II. London: Academic Press.
- ROBINSON, E.M., 1944. A note on strains of tuberculosis from the kudu. Onderstepoort Journal of Veterinary Science and Animal Industry, 19, 23–28.
- ROBINSON, E.M., 1955. Tuberculosis in sheep and goats. Journal of the South African Veterinary Medical Association, 26, 95–104.
- ROBINSON, E.M., 1958. Tuberculosis in pigs in South Africa. Journal of the South African Veterinary Medical Association, 21, 129.
- RUNYON, E.H., 1959. Anonymous mycobacteria in pulmonary disease. Medical Clinics of North America, 43, 273.
- RYAN, T.J., LIVINGSTONE, P.G., BAILEY, J.B., CARTER, C.E., CREWS, K.B. & TIMBS, D.V., 1998. Tuberculosis control in livestock in New Zealand: The current situation and future directions. In: THRUSFIELD, M.V. & GOODALL, E.A., (eds). Society for Veterinary Epidemiology and the Preventative Medicine. 25–27 March, Ennis, Ireland. pp. 1–14.
- SANSON, R.L., 1988. Tuberculosis in goats. Surveillance, 15, 7–8.
- SCHLIESSER, TH., 1976. Vorkommen und Bedeutung von Mykobakterien bei Tieren. Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene, Erste Abteilung. Originali Reihe A. 235, 184–194.
- SCHLIESSER, TH., 1982. Die Rindertuberkulose im Wandel der letzen 100 Jahre (1882–1982). Praxis der Pneumologie, 36, 151–156.
- SCHMIEDEL, A., 1968. Development and present state of bovine tuberculosis in man. Bulletin of the International Union Against Tuberculosis, 40, 5–37.
- SCHMITT, S.M., FITZGERALS, S.D., COOLEY, T.M., BRUNING-FANN, C.S., SULLIVAN, L., BERRY, D., CARLSON, T., MINNIS, R.B., PAYEUR, J.B. & SIKARSKIE, J., 1997. Bovine tuberculosis in free-ranging white-tailed deer from Michigan. Journal of Wildlife Diseases,
- SCHNEIDER, H., 1977. Analyse der Tiergesundheitssituation in Südwestafrika/Namibia—Vergangenheit und Gegenwart. Inaugural Dissertation zur Erlangung des Doktorgrades bei dem Fachbereich Veterinärmedizin und Tierzucht der Justus Liebig-Universität zu Giessen.
- SCHNEIDER, W., DOBBELAER, R., DAM, A., JORGENSEN, J.B., GAYOT, G., AUGIER, J., HAAGSMA, J., REES, W.H.G., LESSLIE, I.W. & HEBERT, C.N., 1979. Collaborative assay of EEC standards for bovine tuberculins. Journal of Biological Standardization, 7, 53–66.
- SEIBERT, F.B., 1944. The chemistry of tuberculin. Chemical Review, 34, 107–127.
- SEIBERT, F.B., FIGUEROSA, E.S. & DUFOUR, E.H., 1955. Isolation, identification and classification of proteins of tuberculin and the tubercle bacillus. American Review of Respiratory Diseases,
- SNYMAN, P.S., 1955. The problem of bovine tuberculosis in the Union of South Africa. Journal of the South African Veterinary Medical Association, 26, 241–245.
- STUART, F.A., MANSER, P.A. & MCINTOSH, F.G., 1988. Tuberculosis in imported red deer (Cervus elaphus). The Veterinary Record, 122, 508– 511.
- SUTHER, D.E., FRANTI, C.E. & PAGE, H.H., 1974. Evaluation of a comparative intradermal tuberculin in California dairy cattle. American Journal of Veterinary Research, 35, 380–397.
- TESSARO, S.V., FORBES, L.B. & TURCOTTE, C., 1990. A survey of brucellosis and tuberculosis in bison in and around Wood Buffalo National Park, Canada. Canadian Veterinary Journal, 31, 174–180.
- THOEN, C.O., ARMBRUST, A.L. & HOPKINS, M.P., 1979. Enzyme-linked immunosorbent assay for detecting antibodies in swine infected with Mycobacterium avium. American Journal of Veterinary Research, 40, 1096.
- THOEN, C.O. & HIMES, E.M., 1986. Pathogenesis of Mycobacterium bovis infection. Progress in Veterinary Microbiological Immunity, 2, 198–214.
- THOEN, C.O., HIMES, E.M. & KARLSON, A.G., 1984. Mycobacterium avium complex. In: kubica, g.p. & wayne, l.g., (eds). The Mycobacteria. A Sourcebook. Part B. New York and Basel: Marcel Dekker, Inc.
- THOEN, C.O., HIMES, E.M., STUMPFF, C.D., PARKS, T.W. & STURKIE, H.N., 1977. Isolation of Mycobacterium bovis from the prepuce of a herd bull. American Journal of Veterinary Research, 38, 877–878.
- THOEN, C.O. & KARLSON, A.G., 1986. Tuberculosis. In: Diseases of Swine, 6th edn. Ames: Iowa State University Press.
- THOEN, C.O., QUINN, W.J., MILLER, L.D., STACKHOUSE, L.L., NEWCOMB, B.F. & FERREL, J.M., 1992. Mycobacterium bovis infection in North American elk (Cervus elaphus). Journal of Veterinary Diagnostic Investigation, 4, 423–427.
- THORBURN, J.A. & THOMAS, A.D., 1940. Tuberculosis in the Cape kudu. Journal of the South African Veterinary Medical Association, 11, 3–10.
- VAN HEERDEN, A. & BRÜCKNER, G.K., 1985. Bovine Tuberculosis; Scheme and Manual for the Control and Combating of Bovine Tuberculosis in South Africa, Division of Veterinary Services, Department of Agriculture and Water Supply, Republic of South Africa. Pretoria: Government Printer.
- VAN WAVEREN, G.M., 1953. Tuberkulose ondersoek by runderen met PPD tuberculines. Tijdschrift voor Diergeneeskunde, 78, 735–760.
- VAN WAVEREN, G.M., 1965. Bovine tuberculosis, tuberculins and tuberculin-tests in the Netherlands. Bulletin Office International des Epizooties, 63, 1395–1401.
- VON ZIEGLER, M., 1953. Die Eutertuberkulose des Rindes. Hirzel Verlag, Leipzig. Reviewed in Schweizer Archiv für Tierheilkunde, 95, 410.
- WAYNE, L.G. & KUBICA, G.P., 1986. Family Mycobacteriaceae. In: sneath, p.h.a., mair, n.s., sharpe, m.e. & holt, j.g., (eds). Bergey’s Manual of Systematic Bacteriology. Vol. II. Baltimore, London, Los Angeles, Sydney: Williams & Wilkins.
- WEBB, G.B., 1936. Clio Medica: Tuberculosis. New York.
- WEBER, A., LUTZ, H. & BAUER, K., 1988. Die derzeitige Bedeutung des Menschen für das Auftreten von Mycobacterium bovis infektionen in Rinderbeständen. Berliner und Münchener Tierärztliche Wochenschrift, 101, 341–344.
- WERNER, E., 1981. Übertragung der Tuberkulose auf eine Rinderherde durch eine Tierpflegerin mit Nierentuberkulose. Monatshefte der Veterinärmedizin, 36, 819–820.
- WIEGESHAUS, E., BALASUBRAMANIAN, V. & SMITH, D.W., 1989. Immunity to tuberculosis from the perspective of pathogenesis. Infection and Immunity, 57, 3671–3676.
- WILKINS, E.G.L., GRIFFITHS, R.J. & ROBERTS, C., 1986. Pulmonary tuberculosis due to Mycobacterium bovis. Thorax, 41, 685–687.
- WINDSOR, R.S., DURRANT, D.S. & BURN, K.J., 1984. Avian tuberculosis in pigs: Mycobacterium intracellulare infection in a breeding herd. The Veterinary Record, 114, 497–500.
- WOOD, P.R., CORNER, L.A., ROTHEL, J.S., BALDOCK, C., JONES, S.L., COUSINS, D.B., MCCORMICK, B.S., FRANCIS, B.R., CREEPER, J. & TWEDDLE, N.E., 1991. Field comparison of the interferon-gamma assay and the intradermal tuberculin test for the diagnosis of bovine tuberculosis. Australian Veterinary Journal, 68, 286–290.
- WOODFORD, M.H., 1982. Tuberculosis in wildlife in the Ruwenzori National Park Uganda (Part I). Tropical Animal Health and Production, 14, 81–88.
- WOODFORD, M.H., 1982. Tuberculosis in wildlife in the Ruwenzori National Park, Uganda (Part II). Tropical Animal Health and Production, 14, 155–160.
- WORTHINGTON, R.W., 1967. Mycobacterial PPD sensitins and the non-specific reactor problem. Onderstepoort Journal of Veterinary Research, 34, 345–438.
- WORTHINGTON, R.W. & KLEEBERG, H.H., 1966. The comparative tuberculin test using Onderstepoort avian and bovine PPD Tuberculin. Journal of the South African Veterinary Medical Association, 37, 177–183.
- YAGER, J.A. & SCOTT, D.W., 1985. The skin and appendages. In: JUBB, K.V.F., KENNEDY, P.C. & PALMER, N., (eds). Pathology of Domestic Animals. 3rd edn. Orlando: Academic Press.
- ZAMBORSZKY, Z., KÖRMENDY, B., TILLIV, D. & TUBOLY, S., 1992. Diagnosis of tuberculosis infection in red deer. Proceedings of the International Conference on Animal tuberculosis in Africa and the Middle East, Cairo.
- ZUMARRAGA, M., PAOLICCHI, F., GARBACCIO, S., GIOFFRE, A., CAIMI, K., BIGI, F., ALITO, A., ROMANO, M.I. & CATASDI. A., 2002. Application of the PCR for the detection of M. bovis in tissue samples. Revista de Medicina Veterinaria, 83, 139–141.




