- Infectious Diseases of Livestock
- Part 3
- Contagious bovine pleuropneumonia
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Contagious bovine pleuropneumonia
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Contagious bovine pleuropneumonia
Previous authors: F THIAUCOURT, J J VAN DER LUGT AND A PROVOST*
Current author:
F T H THIAUCOURT - OIE Expert and Head of CIRAD CBPP Reference Laboratory, Veterinarian, PhD, HDR, TA A117 Campus de Baillarguet, Montpellier, Occitanie, 34398, France
Introduction
Contagious bovine pleuropneumonia (CBPP) is an acute, subacute or chronic disease of cattle and occasionally of water buffaloes (Bubalus bubalis) and yack (Bos gruniensis) caused by Mycoplasma mycoides subsp. mycoides (Mmm). The acute to subacute disease is characterized by serofibrinous pleuropneumonia and severe pleural effusion. Pulmonary sequestra result from the development of chronic, often subclinical, disease.
A recent study based on molecular phylogeny showed that this pathogen probably emerged in Europe in about 1700, i.e. relatively recently, probably from a small ruminant source.34 The disease’s subsequent spread around the world through cattle trade in the 19th Century followed by disappearance from most of these locations, with the notable exception of sub-Saharan Africa, is traced in that paper. Furthermore, due to its persistence in East, West, Central and some parts of southern Africa, it currently exhibits considerable genomic diversity in the sub-Saharan Africa.34
The disease was first unequivocally described in 177344 and has, with the exception of South America and Madagascar, occurred throughout the world at some time. It was eradicated from North America and Australia in the mid-20th Century and probably from Western Europe by the turn of the 21st Century.34 It is an economically important disease in Africa south of the Sahara65, 84 although in southern Africa Botswana, South Africa, Swaziland and most of Namibia are currently recognized as free of the disease (http://www.oie.int/en/animal-health-in-the-world/official-disease-status/cbpp/list-cbbp-free-members/).
The disease was present in some countries of the Near- and Middle East, for example in Kuwait in 1974, but the current situation is uncertain. Pakistan reported its last outbreak in 1997 and India in 1990.84, 95 As of 2017, China, India, Singapore and Australia were recognized as free from CBPP by the OIE (http://www.oie.int/en/animal-health-in-the-world/official-disease-status/cbpp/list-cbbp-free-members/).
Sporadic outbreaks occurred in southern Europe until 1999. The disease was detected in 1967 at the French-Spanish border and then in 1983 in Portugal after an absence of 30 years. Two regions, Entre-Douro e Minho and Beira Litoral, were particularly affected. Intensive test-and-slaughter policies resulted in a dramatic reduction in the number of outbreaks between 1993 and 1999, when the last case was detected. Outbreaks also occurred in Italy in 1990 to 1993 in two regions, Lombardi and Puglia. The disease was eliminated after extensive slaughter-out policies had been adopted but the origin of the outbreaks was never identified.
The presence of CBPP in East, West and Central Africa as well as northern reaches of southern Africa severely restricts rural economic development and the effective utilization of animal resources in those regions. Veterinary regulatory measures aimed at the control the disease hamper effective land-use planning. Prophylactic vaccination campaigns in particular place high demands on the limited budgets of most veterinary administrations confronted by endemic CBPP.
Unlike other parts of Africa, the history of CBPP in southern Africa is well documented. It was introduced into South Africa from The Netherlands by a Friesian bull or bulls landed at Mossel Bay in 1853.79 From there the disease was rapidly disseminated by trek oxen, spreading in all directions along transport routes, and soon reached the northern provinces of the country. Within two years it had killed over 100 000 head of cattle.45 Among its political effects was a major contribution to the Great Xhosa Cattle-killing Movement of 1856 to 1857 that resulted in the starvation of tens of thousands of Xhosa people and the devastation of that nation.79
Namibia was infected in 1856 when a localized outbreak occurred at Warmbad in the south. Although the initial outbreak was contained by quarantine measures adopted by missionaries in the area, the infection reached the central cattle-raising districts in 1859 as a result of cattle introductions from either Botswana or South Africa. High mortalities resulted and the Herero people refer to 1860 as ‘otjipunga’ or ‘the year of the lung’. In 1861 CBPP crossed the Limpopo River to cause heavy losses among the cattle of the Matabele nation in today’s southern Zimbabwe.45
Contagious bovine pleuropneumonia was introduced into Angola from the south, probably by infected cattle belonging to the Dorsland Trekkers who emigrated from South Africa and settled near Humpata in the Huila Province in the early 1880s. Its presence was confirmed there in 1888.69 Extensive use of draught oxen was considered to be the cause of its rapid spread throughout Angola, and by 1914 the whole country had been infected.
The introduction of CBPP into Zambia occurred via Angola. Although eradicated in 1946, CBPP was again introduced into the Western Province of Zambia in late 1969.92 This outbreak resulted in 75 per cent morbidity and caused up to 68 per cent mortality.4 It was eradicated in 1976 but was again reintroduced in 2000 from Angola.
By the time rinderpest appeared in southern Africa in 1896, CBPP was already widespread in the region. The effects of the rinderpest pandemic overshadowed those of CBPP and the disease killed many CBPP-infected cattle. It was not until the turn of the century that the persistence of lungsickness in the cattle populations of southern Africa again presented an obvious problem, and strict control measures were introduced in most countries of the region. These were, however, complicated by other events that occurred about that time, such as the South African (Boer) War, the introduction of East Coast fever, the Herero war in Namibia, and the human influenza pandemic of 1918.45 The disease was eventually eradicated from Zimbabwe in 1904, from South Africa in 1924, and from Botswana in 1939. Botswana was unexpectedly reinfected in early 1995 from northern Namibia.9 Elimination of the disease was achieved in 1997 by the ‘stamping out’ of the entire cattle population of Ngamiland. More than 300 000 cattle were slaughtered and compensated for by the Botswana Government in cash and/or replacement cattle.8 These costly measures were enforced in order to enable the country to restore its CBPP freedom status with the shortest delay.
The disease still persists in northern Namibia91 and in Angola, from where it periodically spreads into the Western Province of Zambia. In Namibia, the high-risk areas are the north-central districts, Kavango and Kunene. In 1997 Namibia commenced massive and regular vaccination campaigns in the north of the country in order to control CBPP. Although CBPP had not been diagnosed in the Zambezi Region (formerly Caprivi) of Namibia since it was eradicated in 1938,11, 68 it was re-introduced into this region in 2003 and controlled after initial blanket antimicrobial treatment followed by regular vaccination.77 In Angola, the disease is endemic throughout the country; this represents a formidable threat for the rest of the region. The reintroduction of the disease into Tanzania in 1990 and its uncontrolled spread throughout the country also poses a severe threat to the southern African region; on several occasions sporadic foci have been reported in north-eastern Zambia. The extent of the disease in the Democratic Republic of Congo (DRC) is uncertain, but according to the OIE Handistatus database for disease occurrence from 1996 to 2004 the DRC suffered two outbreaks in 1997 and the disease was suspected to present from then until 2004 (http://web.oie.int/hs2/sit_pays_mald_pl.asp?c_pays=218&c_mald=7). Since 2005 outbreaks involving at least four districts have been reported to the World Organisation for Animal Health (OIE). (http://www.oie.int/wahis_2/public/wahid.php/Diseaseinformation/statusdetail)
The situation in West and Central Africa in recent years has been characterized by increased numbers of outbreaks and the re-introduction of the disease to The Gambia and Senegal.67 This situation may be explained by the eradication of rinderpest from these regions with the consequent cessation of regular combined vaccination campaigns to control both rinderpest and CBPP.
Recent CBPP vaccination campaigns in Africa have not covered a large proportion of the cattle population. The estimated annual production of CBPP vaccine in Africa over the period 2015 to 2016 was 40-50 million doses (www.oie.int), which represents about 10 per cent of the total cattle population at risk. In addition, low protection rates have been afforded by the vaccine, probably related to factors such as poor handling and administration methods. However, whether it is possible to eradicate CBPP through mass vaccination with the available vaccines, as the sole control measure, is doubtful.103
Aetiology
Mycoplasma mycoides subsp. mycoides (Mmm) is a pleomorphic bacterium that lacks a protecting cell wall, and grows readily under aerobic and anaerobic conditions (see General Introduction: Mollicutes). It belongs to the so-called Mycoplasma mycoides cluster,30 a group of five mycoplasma species or subspecies that are all ruminant pathogens and whose taxonomy was revised in 2009.59 Based on 16S rRNA gene sequence analysis,110 the group seems to be more closely related to spiroplasmas, which are plant pathogens, than to other animal and human mycoplasmal pathogens. The closest relative of Mmm is Mycoplasma mycoides subsp. capri (Mmc), which includes two serotypes “Large Colony” and “capri” that differ according to the type of polysaccharide expressed at their membrane surface.21, 30, 99 Mmc “Large Colony” serotype expresses a capsular polysaccharide (CPS) in common with Mmm. This CPS is highly immunogenic, as the polysaccharide moiety is linked to a lipid moiety anchored in the membrane, and induces the production of IgMs at an early stage of the disease. That explains the cross reactions observed in some serological tests such as complement fixation or agglutination. Mycoplasma mycoides subsp. capri is a common pathogen of small ruminants and causes a ‘contagious agalactia’ syndrome characterized by the development of mastitis, arthritis, keratitis, and, in kids, pleuropneumonia and septicaemia. Some strains may have a specific pulmonary tropism and the disease caused by them has to be clearly differentiated from contagious caprine pleuropneumonia, which is caused by Mycoplasma capricolum subsp. capripneumoniae, the former ‘type F38’54 (see Contagious caprine pleuropneumonia). Differentiation between Mmm, and Mmc was difficult until recently; it can now be achieved by the use of tests based on monoclonal antibodies, polymerase chain reaction (PCR) assays or whole genome sequencing.
Various types of broth can be used to culture the organism.84 (Manual of Diagnostic tests and Vaccines for Terrestrial Animals:OIE www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals). These should contain meat extract or peptone, or both, as the basic medium, as well as fresh extract of baker’s yeast and 10 to 20 per cent serum, preferably from horses. Various additives can be included, such as glucose, sodium pyruvate, DNA, sterol, and antibacterial and antifungal drugs in the case of primary isolation from contaminated samples. Growth is relatively slow; 24 to 48 hours are usually required before growth is visible. Primary isolation may take longer or necessitate subculture, particularly where samples are of inadequate quality. Primary culture in liquid medium often yields the typical aspect of a whitish ‘comet’, a ‘cloud’ floating in the medium that is easily dispersed by agitation. Solid media are prepared by the addition of 1 per cent agar to the fluid medium; Mmm colonies on solid media have the typical ‘fried-egg’ appearance. After 3 to 4 days of incubation, the colony size seldom exceeds 1 mm in diameter and, in order to visualize them, a stereomicroscope needs to be used.
Mycoplasma mycoides subsp. mycoides strains were formerly considered to be homogeneous but recent evidence has shown that genotypes can be distinguished by use of molecular techniques such as restriction analysis of whole DNA,82, 98 southern blotting,25, 60 whole genome sequencing and extended multilocus sequence typing.34 African and European Mmm strains have been differentiated on the basis of genomic differences25, 108 indicating that the outbreaks that occurred in 1967 and 1982 at the French-Spanish border, and in 1980-1999 in Spain, Portugal and Italy were not due to re-introduction from Africa, i.e. they were more likely due to a resurgence of CBPP from a region or regions from which it had not been completely eliminated.
As indicated in the introduction, insight into the evolutionary history of CBPP was obtained through comparison of a representative number of Mmm genomes of European and African origin using phylogenetic and Bayesian analysis for molecular dating.34 The ancestor to all Mmm strains was dated to around 1700 while that of the ancestor to all African strains was 1800, at a time when CBPP was well known to occur in Europe. All the Mmm strains isolated in Europe after 1980 derive from a single clone that possesses an 8.8kb deletion not present in their European ancestor. The strain isolated at the French-Spanish border in 1967 does not possess that deletion, which includes genes involved in glycerol metabolism. The overall estimated mutation rate for the studied set of genes was 5 x 10-7/site/year.
Identification of T1 vaccine strains is possible by PCR.58 However, although genomic variation can be used for molecular marking, the linkage between genomic variation and pathogenicity is not yet established. Within the ‘Mycoplasma mycoides cluster’, Mmm strains have the peculiarity of possessing a number of ‘insertion sequences’ that may account for the greater part of genome variability.25, 109 These multiple copy insertion sequences and the presence of large duplicated genome fragments explain why it took so long to obtain a complete genome sequence for the reference strain Pg1.111 Such duplicated elements may have originated from the recent adaptation of Mmc caprine to enable replication in cattle, accompanied by the acquisition of a novel and transposable element, IS1634, probably derived from another mycoplasma such as M. bovis.101
Mycoplasma mycoides subsp. mycoides is susceptible to environmental factors such as temperature, desiccation or light and, on average, only survives outside the host for up to three days in tropical areas and up to two weeks in temperate zones. Cultures of Mmm can be inactivated by ultraviolet radiation within a few minutes. The organism is inactivated within 60 minutes at 56 °C and within two minutes at 60 °C, but can survive more than 10 years in frozen infected pleural fluid. The organism is inactivated by a 1 per cent phenol solution in three minutes, a 0,05 per cent formaldehyde solution in 30 seconds, and a 0,01 per cent mercuric chloride solution in one minute (Manual of Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals).
The serial passage of Mmm in culture and in animals can alter both virulence and pathogenicity. That provided the basis for the development of attenuated vaccines.
Epidemiology
Mycoplasma mycoides subsp. mycoides primarily affects cattle, water buffaloes (Bubalus bubalis) and yaks (Bos grunniens).55 There are conflicting opinions as to whether there is a difference in susceptibility between Bos indicus and Bos taurus cattle breeds;62 but T1 vaccine is equally efficacious in both bovid species.
The susceptibility of individual animals varies, as it has often been observed that a proportion of cattle are naturally resistant to the disease. Similarly, post-vaccinal reactions occur in varying proportions of cattle herds,86 reflecting higher susceptibility of some animals.
Under natural conditions cattle are the most CBPP susceptible species. Water buffaloes seem less susceptible although typical lesions were observed in that species in Italy.88 In one instance Mmm was isolated from a sequestrum in a water buffalo85 but the role of this species in the transmission of CBPP is considered to be minor. The organism has been isolated from the lungs of naturally infected goats,52 but goats appear to have limited susceptibility judging by an experimental transmission study.118 This observation is supported by the fact that CBPP was eradicated from Botswana after the outbreak there in 1995 by slaughter of only the cattle population, i.e. the goat and sheep populations were not ‘stamped out’. Previous reports of detection of CBPP in wild animals by serology are probably incorrect because the technique used (complement fixation) is prone to cross-reaction with antibodies against related mycoplasmas such as Mmc. Two African buffaloes (Syncerus caffer) were inoculated experimentally with Mmm and both died after having shown high complement fixing antibody levels to the organism in their sera. Mycoplasma mycoides subsp. mycoides was isolated from lungs, liver and spleen in pure culture from one of them but neither had any lesions, which casts doubt on the actual cause of death.94
Three factors are of primary significance in the transmission of CBPP: intimacy of contact between infected and susceptible animals, the rate of excretion on the part of infected animals and the size and density of the susceptible cattle population. The estimated mean incubation period is about 4 to 6 weeks.65, 107
Natural transmission of CBPP occurs by droplet infection derived from clinically diseased cattle or from carriers excreting the organism to susceptible animals in close contact.49 Mycoplasma mycoides subsp. mycoides occurs in bronchial secretions, nasal discharges and exhaled air of diseased animals. Dissemination of infection occurs most readily among closely stabled or trucked animals. Aerosols containing infected droplets may spread the disease over distances of 20 m or more. Such transmission was suspected in Italy during the 1993 outbreaks,40 and experimental studies with other Mycoplasma species have shown that live organisms could be retrieved over much longer distances, in the range of 5 to 10 km.78 Even under extensive sub-Saharan African farming conditions spread of the disease may be rapid, infection being facilitated by the general practice of kraaling animals at night and congregation of large numbers of animals at places such as watering troughs or markets.
Transmission occurs most commonly by direct contact between susceptible and diseased animals. The infection is not transmitted to animals introduced into crushes, stock sales yards or transport vehicles that have previously been occupied by infected cattle. Neither ingestion of infected fodder nor direct exposure to diseased organs of animals suffering from CBPP results in transmission.49
Mycoplasma mycoides subsp. mycoides may be present in the urine of cattle affected with severe disease; a ‘urinary tract to nose’ route of transmission or the spread of infection through aerosols of urine droplets thus appears to be possible.66 The organism was also isolated41 or detected by PCR in bull semen.98 Therefore, direct sexual transmission and indirect transmission by frozen semen or embryos might occur; these mechanisms of transmission, however, require further investigation.
Apart from clinical cases of the disease, there are a number of other ways in which the infection can be introduced or maintained. Mycoplasma mycoides subsp. mycoides may be present in the nasal passages of cattle for 40 days during the incubation period of the disease, before any serological response to it can be detected.50 Such animals may thus disseminate the infection to susceptible animals; they are suspected to play an important role in the spread of the disease in both commercial and smallholder cattle farms. Another way in which infection may be introduced or maintained has long been proposed. During the course of the disease, pulmonary sequestra may develop in affected animals and these may persist in clinically recovered cattle for many months. It is difficult to obtain estimates of the mycoplasma load that can be excreted by these animals. The sequestra are comprised of necrotic tissue that becomes encapsulated by fibrous connective tissue and may harbour viable mycoplasmas for 12 months or longer. Such chronically affected animals are known as ‘lungers’.45 The role of these lesions in the persistence and transmission of CBPP is, however, debatable.73 Completely closed capsules are not likely to play any role in the dissemination of infection, but rupture of the capsule or reactivation of lesions in the surrounding lung tissue may lead to transmission of CBPP to susceptible animals. Under such circumstances, clinically recovered animals harbouring such lesions could be responsible for spreading the disease.
Infected young calves usually develop arthritis particularly of the carpus and tarsus, and until relatively recently it was thought that they did not develop pleuropneumonia. In the 1995 epidemic in Botswana, however, severe CBPP lesions were seen in the lungs of calves as young as three to six months of age.29 In view of this it is hypothesized that in endemic situations calves in the first few months of life are partially protected by progressively waning colostral immunity and therefore do not develop typical disease, whereas those in an epidemic situation do not have this protection and, as a consequence, develop fully-fledged disease.
Within a herd that is infected, four groups of animals can be distinguished: those that are naturally resistant (15 per cent); those that may harbour the mycoplasma without any overt sign of disease (15 per cent) and may undergo sero-conversion; those that are moderately susceptible (55 per cent) and able to control the lung inflammation leading to sequestrum formation of various sizes; and those that are not able to control the lung inflammation (15 per cent) and die. The proportions of these four groups may vary significantly from one outbreak to another and this probably depends on the virulence of the Mmm strain and also factors such as breed48 or any event that may impact the immune response of the animal. Hudson and Turner proposed a scoring system to quantify and integrate the various criteria of infection and susceptibility in order to evaluate the protection afforded by vaccines.50 These scores varied amongst naïve groups of experimentally infected cattle. This reflects what is observed in the field, where the outcome of the disease may be modified by administration of antimicrobials.
The level of susceptibility of animals in a given herd varies considerably between individuals. Ten to 60 per cent of cattle may apparently be resistant, and as few as eight per cent of a herd may develop clinical signs. Survivors of an outbreak develop protective immunity. Thus, in a closed-herd situation, even under African conditions, there is a tendency for the disease to “disappear”, albeit over a long period of time. In this situation new-born calves or naïve introductions constitute a nucleus of susceptible animals. In Africa, movement of animals between herds is common and is the direct cause of many outbreaks. This can occur through market transactions, dowry payments, cattle congregation at water points and pastures, or exchange of animals for reproduction or ploughing. It can also occur illegally through the marketing of sick animals in order to escape control measures, or by theft. Civil strife is a major cause of CBPP transmission as it reduces the effects of control measures and also results in uncontrolled animal movements.
The basic reproductive number (R0) of CBPP has been estimated to range between 1.24 and 256 or from 3.2 to 4.61 These values are relatively low compared to many epidemic viral diseases and this has practical implications. For example, it implies that herd immunity of 50 to 75 per cent would be sufficient to interrupt transmission.10
Pathogenesis
The pathogenesis of lungsickness is poorly understood as its investigation has been hampered by the difficulty and cost of reproduction of CBPP in cattle and the absence of a reliable laboratory animal model.
Infection via the respiratory tract causes bronchiolitis and pneumonia.49 In this process Mmm attaches to the surface of epithelial cells of the respiratory system and is thus prevented from being eliminated in mucus secretions. Binding to adult bovine lung epithelial cells is greater than for foetal lung epithelial cells or for endothelial cells and fibroblasts.13 This cellular adhesion, in which the surface of the mucous membrane is colonized, is central to the pathogenicity of the disease. In contrast to other Mycoplasma spp., such as M. pneumoniae, there is no proof as yet that attachment occurs as a result of the production of an adhesin by the organism. The attenuation of Mmm by in vitro passage is proof that virulence genes do exist, and their existence has recently been confirmed by comparative genomics.57 These genes may not be a necessity for the growth of Mmm and are capable of undergoing random mutation in vitro, thus altering their expression in vivo.
Multiplication of Mmm in the lung results in severe local inflammation. The type of antigen that triggers this inflammation is not known. The types of cells and cytokines involved require further study, but results of a recent experimental infection study have identified increased levels of alpha tumour necrosis factor (αTNF) and interleukins IL-1β and IL-17A associated with acute lesions in infected lungs, as well as increased numbers of macrophages and neutrophils.97 The lesions of CBPP in naturally infected cattle may be restricted to lung tissue simply because of its mode of transmission, as experimental inoculation may result in the development of lesions at the site of inoculation and in other organs but not in the development of lesions typical of the natural disease.112 Subcutaneous inoculation of a pathogenic Mmm strain results, after 10 to 25 days, in local oedema known as a ‘Willems reaction’, the term being derived from the name the person who conducted inoculation trials performed in 1852.112
The pseudo-capsule of Mmm organisms is composed of the carbohydrate galactan, a bêta(1>6)-galactofuranose homopolymer. This galactan is present in two forms, the CPS in which galactane is linked to a lipid moiety anchored in the membrane and the excreted polysaccharide (EPS) composed of pure galactan, which is secreted outside the mycoplasma body (either in vivo or in vitro). CPS is a potent antigen that will induce the production of antibodies, notably IgMs, which recognize both CPS and EPS. CPS is a potential determinant of virulence because it appears to play a role in attachment to target tissues.37 It may also contribute to the resistance of the organism to phagocytosis38 and induce the formation of autoantibodies to pneumogalactan. The membrane-bound galactan, which is linked to a lipid moiety, may play a role in the stimulation of the inflammatory reaction. On the other hand, EPS has anti-inflammatory effects.104 The complete genome sequence of Mmm Pg1111 failed to reveal obvious virulence factors such as toxins, which could be due to the fact that Pg1 is no longer virulent. Subsequently published Mmm genome sequences derived from virulent strains such as “Gladysdale”,39, 114 did not add clues to virulence factors. Differences in pathogenicity between European and African strains have been postulated but it is difficult to compare pathogenicity between the differing environmental circumstances that prevail in Africa and Europe. The absence of a glycerol transporter gene in the most recent European strains might account for the possible lower virulence through lowered production of hydrogen peroxide.81, 107 However, vaccine strains, such as T1, and the European strain PO-67, isolated in 1967, possess this glycerol transporter gene.

Other factors may play a role in the pathogenesis of the disease; for example, it is thought that external and internal parasite infestations may have immuno-modulating effects, leading to lower susceptibility, or that other intercurrent diseases impair normal immune responses. It is possible that animals in better physical condition exhibit increased susceptibility to CBPP. A better insight into the pathogenesis of the disease will be obtained once the roles of specific types of cells and the cytokines produced during the inflammatory process have been fully elucidated.
In the acute stage of CBPP, antibodies are produced in high quantities. IgMs that recognize both CPS and EPS may be responsible for the formation of antigen-antibody complexes and local renal infarcts that are sometimes observed in infected animals. The course of the disease does not seem to be influenced by the presence of antibodies directed towards surface proteins, with sequestra being formed in spite of high antibody titres.90 Some Mycoplasma species, including Mmm and those of the M. mycoides cluster, have the unique ability to bind and cleave immunoglobulins,12 which could provide a mechanism for escaping the host’s humoral immune response.
Clinical signs
The clinical signs in cattle have been reviewed in a number of publications,31, 35, 49, 84 but are mostly attributable to pulmonary pathology.
Under natural conditions the incubation period is rarely less than three to six weeks and may exceed three months. However, following massive aerosol exposure, the incubation period may be as short as 15 days. As a rule of thumb, cattle that have been in contact with animals suffering from CBPP should be considered as potentially infectious for at least six months after exposure.
Clinical disease may be acute, subacute, or chronic. In the initial stages of an epidemic the disease tends to be acute, but as the epidemic progresses sub-acute and chronic cases predominate. In endemic areas the disease tends to be mostly chronic and subclinical.


Acutely affected animals have an elevated body temperature, are listless, and manifest signs of respiratory distress. They often grunt as if suffering from pain and occasionally a soft, moist cough is audible, which is aggravated by and becomes more frequent on exercise or on percussion of the chest. As the disease progresses, coughing increases both in frequency and in intensity, the animal is reluctant to move, and stands with its head extended, mouth open, tongue protruding, and the ‘elbows’ turned out (Figure 1). Contraction of the muscle of the abdominal wall occurs after each inspiration, while expiration is frequently followed by a characteristic grunt or groan. There is usually a mucoid discharge from the nostrils and frothy saliva accumulates around the mouth. The affected areas in the lungs are indicated by the presence of dull sounds on percussion. During the later stages of the disease the animals may develop a mucopurulent nasal discharge (Figure 2), subcutaneous oedema of the lower parts of the chest and abdomen, and emaciation. In some affected animals dyspnoea is aggravated by the presence of large volumes of exudate in the thorax.
In subacute cases the lung lesions are more localized and infrequent coughing may be the only clinical sign. Usually the lesions resolve without the disease being noticed clinically.
The only clinical signs in chronic cases are emaciation and a cough, which commonly occurs when the animal rises.
In calves up to the age of six months the only sign of disease is usually arthritis, particularly of the carpal and tarsal joints. Pulmonary lesions may or may not develop (see Epidemiology).
Once clinical signs are evident, affected animals may die within two to three weeks; acutely affected animals, however, may die within one week as a result of the development of severe serofibrinous pleuropneumonia and/or a massive pulmonary infarct. In many cases the clinical signs gradually disappear, after which the general condition of affected animals improves rapidly. Severely affected animals that do not die may take many months to recover. Recovered cattle may harbour sequestra in their lungs in which Mmm may remain viable for more than 12 months (see Epidemiology).
In water buffaloes, clinical signs were absent on a farm where five of 1 967 animals in contact with 2 539 cattle had complement fixing antibodies to the agent in their serum. Only one buffalo had a lung lesion, consisting of a typical sequestrum, from which Mmm was isolated.88
Pathology
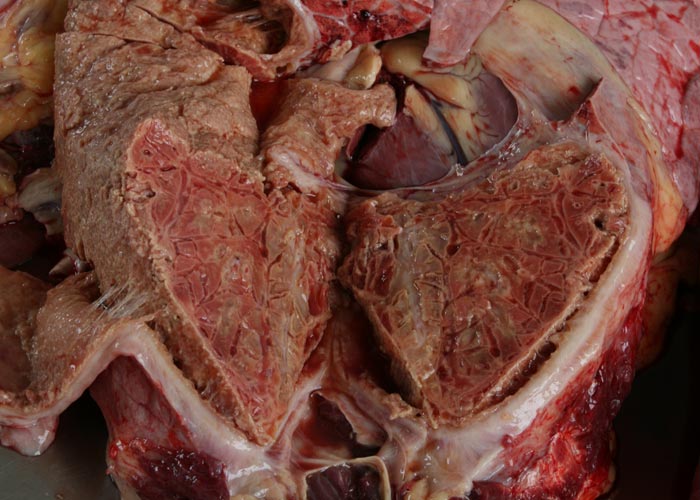
During the acute stage of the disease, uni- or bilateral pleural effusion that may exceed 10 litres is present (Figure 3). The exudate is yellow to yellowish-grey, clear or turbid, contains pieces of fibrin, and is generally accompanied by pronounced localized or diffuse fibrinous parietal and visceral pleuritis (Figures 4 and 5). During the early stages of the disease, multiple well-circumscribed areas of consolidation are scattered throughout the lungs in some animals. Well-developed lesions manifested as severe fibrinous pleuropneumonia that tends to be unilateral, involving a portion of a lobe, an entire lobe or more than one lobe (Figure 6).
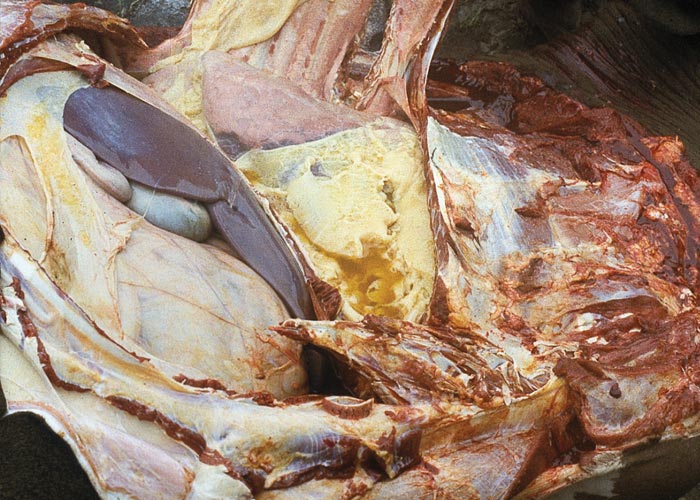

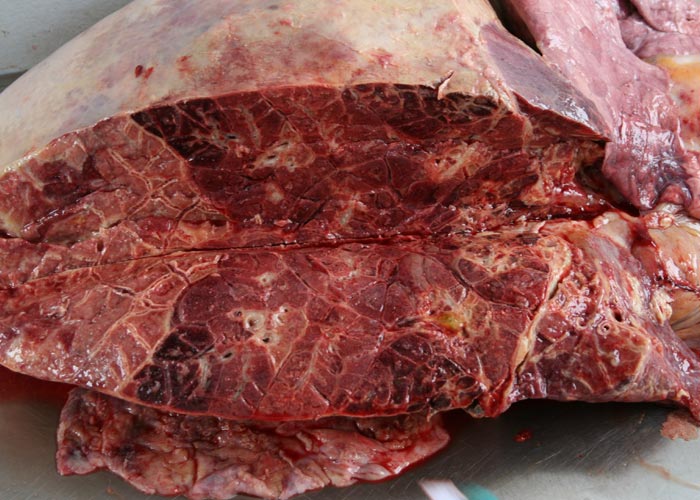
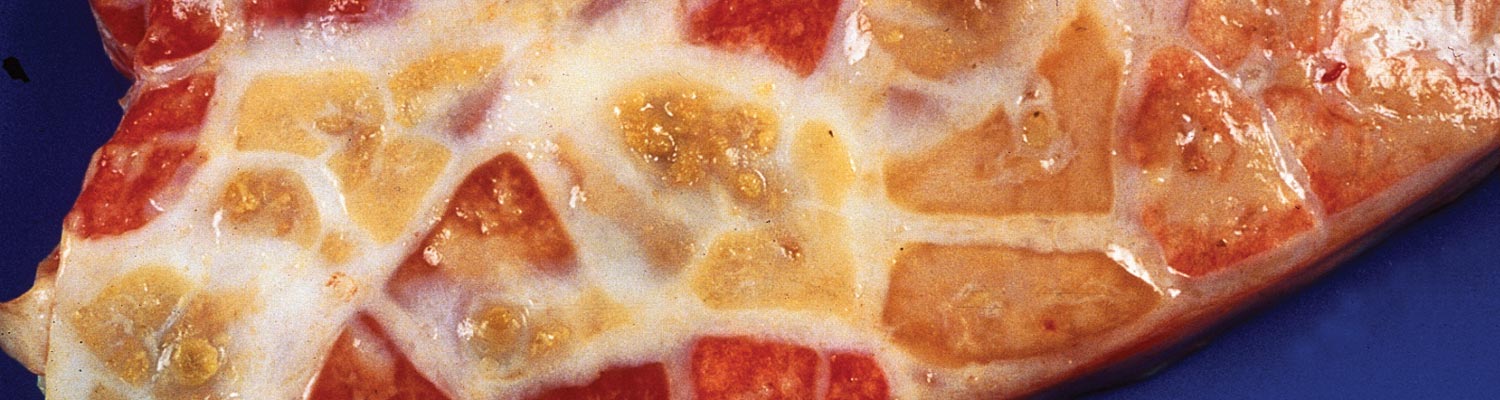
Figure 6 ‘Marbling’ of the lung as a result of different stages (red and grey hepatization) of the pneumonic process and distension and ‘beading’ of interlobular septa (Courtesy of Dr Massimo Scacchia, Istituto Zooprofilattico Sperimentale dell’Abruzzo e del Molise "G.Caporale" Via Campo Boario, 64100 Teramo, Italia)

The pneumonic areas are characterized by ‘marbling’ caused by the juxtaposition of lobules that show varying stages of grey or red hepatization and relatively unaffected lobules (Figures 6 and 7). The ‘marbling’ is accentuated by the distension of interlobular septa and the interstitium surrounding blood vessels and airways by serofibrinous, sometimes blood-stained, exudates (Figure 8). Copious amounts of a clear, yellowish fluid exude from the cut surface of acutely affected lungs. The walls of the bronchi are thickened by oedema, and their lumens often contain fibrinous material. The interlobular septa and the parenchyma bordering areas of pneumonia may be oedematous. Multiple thrombi occur along the course of lymphatic vessels in interlobular septa with consequent irregular lymphangiectasis. The latter often imparts a ‘beaded’ appearance to the septa (Figures 7 and 9). Infarcts, varying in size from about 10 to 300 mm, are frequently present in affected lung tissue (Figure 9).
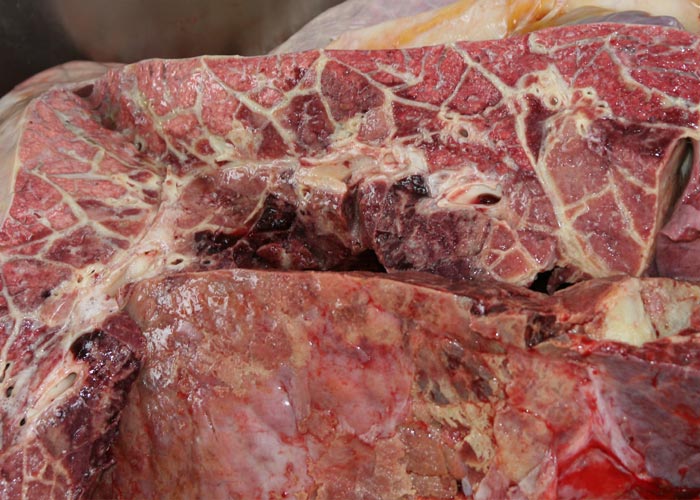
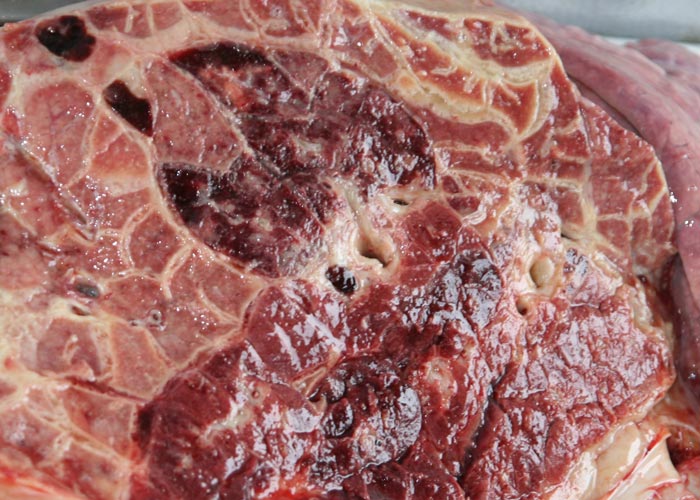
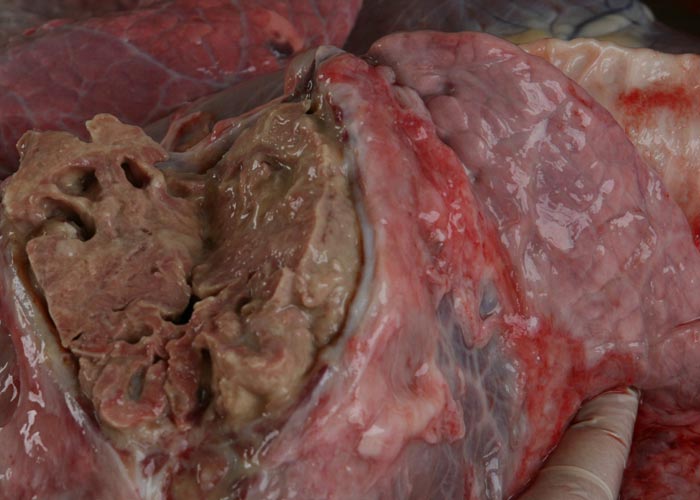
Sequestra (see Figures 10 to 15) result from thrombosis of inter- or intra-lobular arteries and are manifested as well-demarcated areas of necrosis separated from the surrounding tissue by irregular, yellowish-grey reaction zones. The infarcts subsequently become sequestered from the adjacent parenchyma by granulation tissue.
At necropsy, cattle that die of chronic lungsickness are usually emaciated, with lesions evident in the lungs and thoracic cavity.23, 45, 49
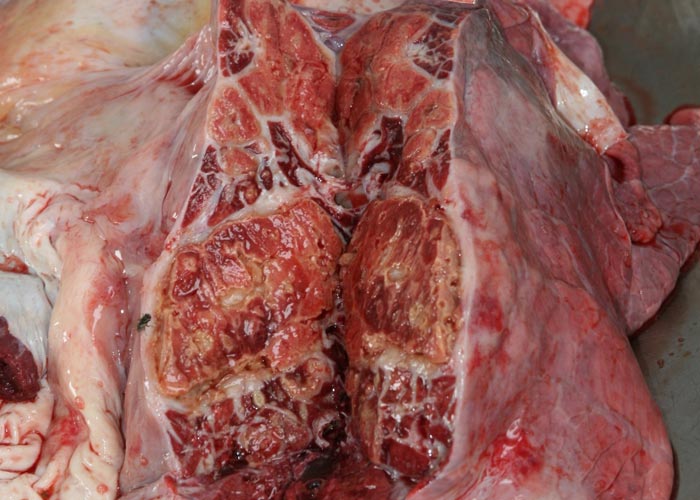
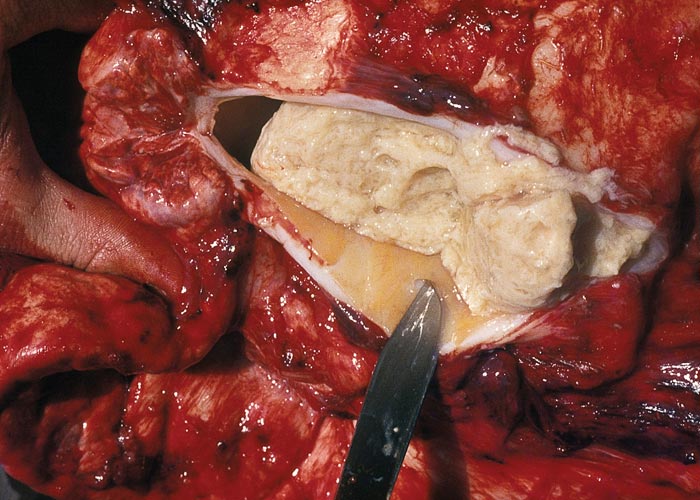
In chronic cases one or more sequestra, which may vary from less than 10 to 300 mm in diameter and involve more than one lobe, are usually present in the lungs. In some animals, however, they may be very large and even involve a whole lobe. Over a period of three to four months smaller sequestra become replaced by fibrotic scars,23 while the larger ones often persist for 12 months or longer. Recently formed sequestra are surrounded by a thin fibrous capsule (see Figure 12), the necrotic content is firm, pinkish to yellowish-grey, and the original pulmonary architecture may still be discernible to some extent. In longer-standing cases the connective tissue thickens and a thin layer of a yellowish, semi-solid material may separate the firmer necrotic contents from the wall. In such sequestra the contents ‘shell out’ easily (see Figures 13 to 15). Secondary bacterial infection of sequestra occurs frequently, resulting in the content being purulent and liquefied. In some cases rupture of the capsule enables the content to spill out into adjacent bronchi.

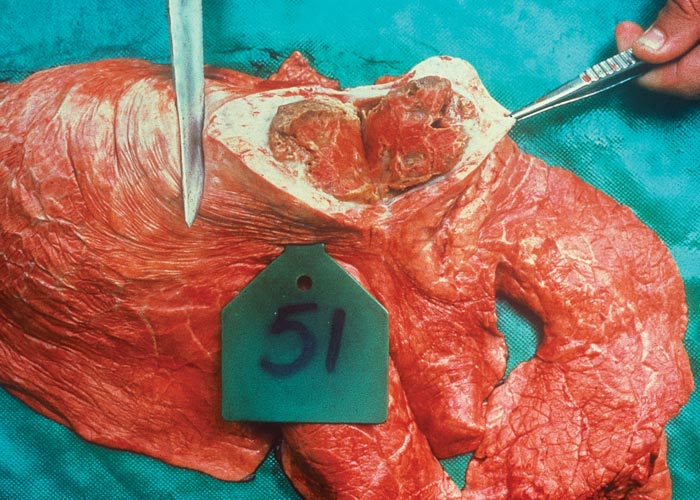
In animals that have completely recovered, a process that may take many months, former infarcts are represented by fibrous scars, and areas of scar tissue are present in the healed lung and pleura. The latter is often thickened with fibrous adhesions remaining between the visceral and parietal pleural surfaces.
Lesions in other organs are often seen in acute cases of lungsickness. The mediastinal and bronchial lymph nodes may be markedly oedematous, while necrotic foci of up to 20 mm in diameter may also be present in these nodes.49 In many cases serofibrinous pericarditis with copious amounts of exudate occurs.
Infarcts, which are generally multiple, small and whitish are not uncommon in the kidneys,23, 49, 64 where they probably result from occlusion of the renal arteries by septic emboli originating in the lungs.
Serofibrinous polyarthritis and tenosynovitis, affecting particularly the carpus and tarsus, are common lesions in infected calves.49, 106 The joints may remain swollen and thickened months after infection as a result of severe fibrosis of the joint capsule. Calves may also develop necro-granulomatous valvular endocarditis and myocarditis.105 Myocardial lesions occur mainly in the atria.
‘Occult’ cases of CBPP, in which only the mediastinal lymph nodes are oedematous with no discernible lung lesions, have been reported.45, 116 The causative organism can usually be isolated from these lymph nodes.
Microscopically, a characteristic feature is the presence of pneumonic foci in a variety of developmental stages in adjacent alveoli or lobules.
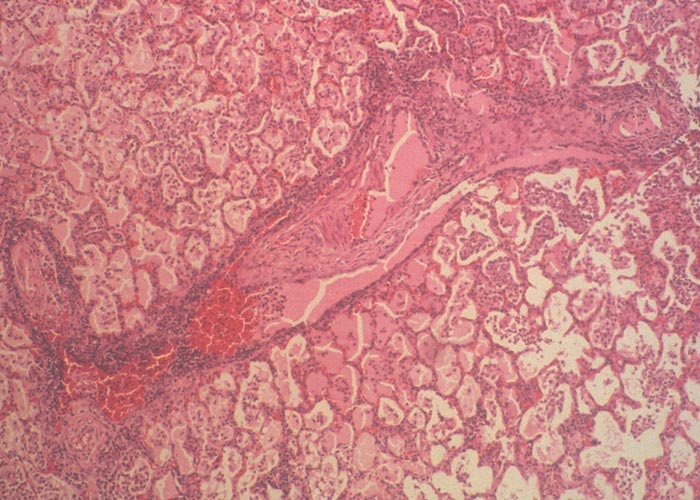
Initially severe hyperaemia of the alveolar capillaries occurs, following which the alveoli become flooded with serofibrinous exudate containing variable numbers of neutrophils, macrophages and red blood cells. As the lesions progress, the amount of fibrin increases, and in some areas numerous necrotic neutrophils fill the alveoli (Figure 16). The interlobular septa are widened by the accumulation of copious amounts of serofibrinous exudate and irregularly distended lymphatic vessels. Necrotic neutrophils collect along the inner walls of lymphatic vessels, particularly along the margins of the septa, where distinct inflammatory zones are frequently discernible. A fibrinopurulent exudate similar to that in the alveoli is often found in and around bronchi and bronchioles of affected lobules. The bronchial mucosa and submucosa are oedematous and infiltrated by neutrophils and macrophages.
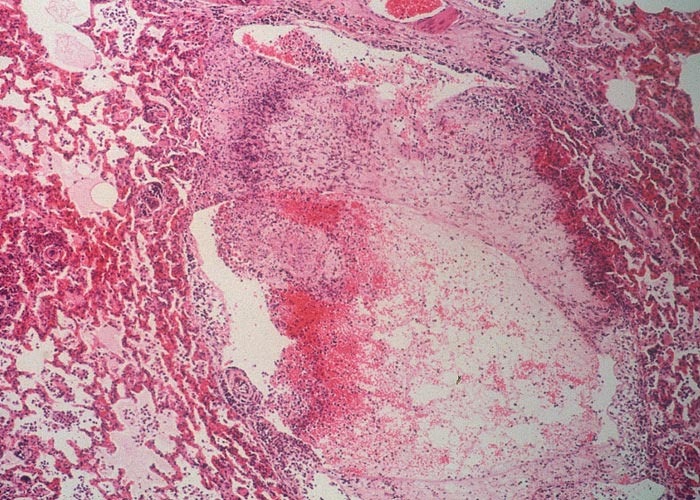
The peribronchial and perivascular areas, and especially the lymphatic vessels, are widely distended by an exudate similar to that of the septa. As the lesions progress, these areas and the septa become thickened by fibrous connective tissue and by infiltrations of lymphocytes and plasma cells. In some parts of the lung the alveolar walls contain varying numbers of round cells and evidence of fibroplasia.
Vasculitis and thrombosis of intralobular and interlobular arteries and the lymphatics are frequently present in pneumonic parts (Figure 17). Involvement of the arteries culminates in necrosis of a lobule or part of a lobule, or in infarction of multiple lobules and intervening septa.34 Initially a zone of necrotic inflammatory cells separates the infarcted areas from the surrounding tissue, but later granulation tissue followed by fibrosis at the edges of these lesions sequestrates the necrotic tissue from the neighbouring parenchyma. Follicle-like aggregates of lymphocytes may be evident in the fibrous capsule of some of the sequestra, as well as in the peribronchial and perivascular areas.
Microscopically, the lesions in lymph nodes are characterized by hyperplasia of lymphoid tissue in the follicles and medullary cords; accumulation of oedematous fluid, fibrin and neutrophils in the subcapsular, cortical and medullary sinuses and trabeculae; vasculitis and thrombosis of blood vessels and lymphatics in the trabeculae; and necrosis of the lymphoid tissue.34, 49
Joint lesions in acute cases consist of hyperaemia, oedema and infiltration of predominantly lymphocytes and macrophages into the synovial membranes. Thrombosis of lymphatics and blood vessels and fibrin deposition on the synovial surfaces also occur. In chronic cases the joint capsule is thickened by connective tissue and hypertrophy of the villi of the synovium in some areas.106
Diagnosis
It is impossible to diagnose CBPP on clinical signs alone as the initial signs may be slight or indistinguishable from severe pneumonia caused by a range of other infectious and parasitic organisms. Therefore pathological, microbiological, molecular and/or serological diagnostic methods should be applied in order to reach a definitive diagnosis. As the pulmonary lesions of CBPP are distinctive, post-mortem lung examination at abattoirs provides a practical way of identifying cases during outbreaks.
New outbreaks can usually be traced to the introduction of cattle into a herd or interaction of herds with cattle newly introduced to the locality. It is therefore essential to obtain a detailed history of the herd.
For the laboratory diagnosis of CBPP the quality of the samples submitted is of primary importance. That holds true even though the isolation and identification of Mmm is now simpler than formerly because of the availability of molecular techniques.76 In live animals, nasal swabs, broncho-alveolar washings or pleural fluid – the sample of choice – obtained by puncturing the lower part of the thoracic cavity between the seventh and eighth ribs, provide good diagnostic material. Synovial fluid can also be collected from those animals with arthritis since the organism is recoverable from that site. Nasal swabs should be placed in a transport medium to protect the mycoplasmas and prevent proliferation of secondary bacteria (Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals).
At necropsy one millilitre of pleural exudate is sufficient but it needs to be collected aseptically. Ampicillin can be added to the sample to prevent multiplication of contaminants. Blocks of pneumonic lung tissue (100 mm3 in size) can also be taken for laboratory investigation as well as tissue specimens from the lining of sequestra. Specimens from the mediastinal and bronchial lymph nodes should also be collected. After collection, all samples must be kept at 4°C and delivered to the laboratory within 24 hours. For longer periods they should be frozen at or below -20°C (Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals).
Small pieces of affected lung should also be collected in 10 per cent buffered formalin for histopathological and/or immunohistological examination.
In vitro culture and biochemical and immunological identification tests
The presence of the pathogen varies greatly with the stage of development of lesions; therefore negative results are not necessarily conclusive, particularly in animals treated with antibiotics.
Several media compositions can be used to culture Mmm (Manual of Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals). Pure cultures may be obtained from test material, but where that is not the case classic mycoplasmal identification procedures that include prior cloning steps may be required for accurate identification. In the past, identification was based on biochemical tests such as glucose fermentation (+), arginine hydrolysis (−), reduction of tetrazolium salts (+) and absence of phosphatase activity. In addition, growth inhibition tests using hyperimmune serum against Mmm were performed. These tests, however, are not specific and therefore do not enable clear differentiation from related subspecies of the ‘mycoides cluster’, e.g. Mycoplasma mycoides capri (Mmc). Biochemical methods are described in the OIE Manual of Diagnostic tests and Vaccines for Terrestrial Animals (Manual of Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals).
Molecular identification and typing assays
In disease-free areas the specific and rapid identification of Mmm is of the utmost importance. This can be achieved preferably by PCR but other assays such as immunobinding assays32 and growth inhibition tests53 using specific monoclonal antibodies could also be used. Detection of Mmm can also be achieved using formalin-fixed tissues by immunohistochemistry.18, 89
Various PCR systems have been developed,19, 22, 70, 80 although no single method is preferred. A method based on modification of an assay developed by16 that has been found useful is described in the OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Manual of Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals.
Subtyping of Mmm isolates using molecular methods can be performed by multilocus sequence analysis of relatively low numbers of genes117 or using an extended system.34
Serological tests
Routine serological tests such as complement fixation (CFT) and competitive enzyme-linked immune-sorbent assays (cELISAs), recommended by the OIE, are valid only at the herd level. These tests will naturally not detect animals in the incubation period or late chronic stage of the disease. The CFT lacks specificity and its predictive value may be low in herds where the prevalence is low. In that case Western blotting may be a good choice to confirm the results. Besides, CFT detects mostly IgMs and will yield positive results only for a short period of time of three to four months. The cELISA with monoclonal 117/5 has been designed to yield a very high specificity of more than 99,9 per cent when it is run in a quality- assured environment. It detects antibodies for a longer period than the CFT.
Immuno-blotting provides more specific results but is unsuitable for testing of large numbers of animals (Manual of Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals.
The sensitivity and specificity of the CFT have been estimated at 63.8 per cent and 98 per cent respectively,20 the low sensitivity being due to failure to detect animals in the early (latent) and late (chronic) stages of infection. Despite this problem, the CFT is a useful test because is able to detect infection within eight days and remains positive for only five to six months on average. Furthermore, it is useful for differentiating between serological responses to infection and vaccination with live attenuated vaccine because, following vaccination, CF antibodies persist for only three months at most. Therefore, for animals that have been vaccinated three or more months previously, a positive reaction is indicative of infection. The CFT can therefore be used to monitor the prevalence of CBPP in vaccinated populations. The test, however, is difficult to standardize between laboratories, especially if antigens from different sources are used.
The cELISA is able to detect antibodies to CBPP for longer than the CFT and therefore to reveal the presence of animals with lung sequestra.71 Assessment of this test in both Europe and Africa showed that i) the true specificity of the cELISA was at least 99.9 percent; ii) the sensitivity of the cELISA was similar to that of the CFT; iii) antibodies were detectable by the cELISA in an infected herd very soon after detection by the CFT; iv) and cELISA antibody persists for a longer period of time.9, 53, 74 To enhance its repeatability and robustness, this cELISA is now provided as a ready-made kit that contains all the necessary reagents, including precoated plates (Manual of Diagnostic tests and Vaccines for Terrestrial Animals:OIE - www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals).
As a consequence, the CFT is well suited to detect acute cases while the cELISA is better suited to determine prevalence rates. For both tests prevalence or incidence rates should be established at the herd level ( epidemiological unit). Accordingly, the sampling frames should aim and to include a random selection of herds and targeting those animals that show clinical signs and more chronic cases to increase the sensitivity of detection.
As is the case with the CFT, the cELISA does not detect antibodies in vaccinated animals, thus allowing prevalence studies in vaccinated areas to be undertaken. It also does not detect all infected animals.
A rapid cow-side diagnostic test was developed as long ago as 1954.79 It is based on an inactivated, stained Mmm antigen that can be used in an agglutination test. This test is probably less sensitive and specific than CFT and cELISA but has proven useful for the rapid diagnosis of acute cases in remote areas.
Differential diagnosis
Acute CBPP needs to be differentiated from pneumonic pasteurellosis, in which Mannheimia haemolytica and (less commonly) Pasteurella multocida play major causative roles, as well as haemorrhagic septicaemia caused by Pasteurella multocida types B and E. Haemorrhagic septicaemia is often characterized by swellings of the laryngeal region and brisket, and has an acute to hyperacute course. In acute pneumonic pasteurellosis serofibrinous pleural exudates and fibrinous pleuropneumonia are present. Smears from the affected lung or, in some cases, blood smears stained with Giemsa, Leishman or methylene blue will reveal typical bipolar organisms.
Acute serofibrinous pleuropneumonia that results from inhaling foreign material into the lungs often resembles CBPP closely.
The sequestra in chronic cases of CBPP need to be differentiated from lung abscesses caused by pyogenic bacteria. Culture and histopathology examination are helpful in that regard.
The clinical signs of CBPP and East Coast fever49 and Corridor disease, in which there is marked pulmonary oedema may be confused with those of CBPP. Bovine herpesvirus 1 infection (infectious bovine rhinotracheitis) generally causes lesions in the upper respiratory tract, but in some animals’ bronchopneumonia may develop, characterized by a necrotizing bronchitis and atelectasis, often complicated by secondary bacterial infection with Pasteurella spp.
Control/eradication
There are three methods that can be used to control CBPP: slaughter of infected animals, antibiotic treatment and vaccination. Control of animal movement is a further important element in the management of all contagious diseases. These methods can be applied in various combinations to develop strategies for specific epidemiological situations, animal husbandry systems, and the availability and effectiveness of veterinary services in individual countries or zones.
There are numerous historical examples of countries or continents that have successfully eradicated CBPP. A strict slaughter protocol was used in the USA and the UK at the end of the 19th century to achieve this goal. This was a time when no modern diagnostic tool was available and herd infection status was determined solely by clinical signs and pathology.31 Australia also succeeded in eradicating CBPP in 197328 after more than a decade of intensive effort. Australia’s initial strategy was to lower the overall prevalence of CBPP by ‘cleaning’ infected herds. This was achieved by successive rounds of serological testing (CFT) and elimination of positive animals, accompanied by vaccination of all the other animals using the V5 vaccine strain administered at the tip of the tail. This procedure was repeated every three months as live vaccines do not induce long-lasting immunity. A herd was considered ‘cleaned’ after testing negative twice consecutively. Later in the campaign vaccination was discontinued and complete eradication obtained by slaughter of the remaining infected herds. The strategy was successful primarily because strict control of animal movements was implemented. Animals to be slaughtered in clean areas were transported to abattoirs by sealed trains. More recently, Italy and Portugal regained CBPP freedom by applying a strict herd slaughter strategy after serological detection of positive herds using CFT.85 This eventually resulted in Portugal eradicating the disease in 1999.
In Africa, two situations prevail: countries that are free of CBPP and those where CBPP is endemic. In countries free of the disease, such as the southern part of the Southern African Development Community, the objective is to maintain that status through an effective ban of importation of live animals from countries where CBPP is present. Official trade is easy to control but illegal movements of animals may constitute the main risk of introduction. Where the disease is endemic control strategies need to be implemented and the strategy and measures clearly documented.
Among the tools that can be used to manage CBPP, the use of antimicrobials has long been a contentious issue. Their use was formerly not recommended because of the possible induction of carriers and the emergence of resistant Mmm strains. In addition, it was considered that antimicrobials would suppress the development of clinical signs that, in turn, would delay the recognition of the disease and the implementation of more effective control measures such as stamping out. In fact, very few trials were performed to substantiate the real effect of antimicrobial treatment or to evaluate what role they could play in holistic control strategies. Because they do not have a cell wall, mycoplasmas are naturally resistant to penicillins. They also rapidly become resistant to aminoglycosides. These antibiotics should therefore not be used for the treatment of CBPP. Chloramphenicol should not be used in animals as it is restricted to use in humans. Antimicrobials of the tetracycline family are regularly used for the treatment of CBPP in Africa, but others that are used are macrolides, lincosamines and fluoroquinones that are active against Mmm.14, 22 Tetracycline was efficient in alleviating the clinical signs in clinically affected animals75 and reduced dramatically the lesion score in treated animals, although it did not prevent the persistence of Mmm in treated animals. Conversely, fluoroquinone treatment did not reduce the lesion score or prevent the persistence of Mmm in treated animals, but treated animals failed to transmit CBPP to naïve contacts.47 There are few reports based on field experience. Among various antibiotic treatments that were tried in a field outbreak in Ethiopia, only tylosin prevented the occurrence of relapses while tetracycline or penicillin-streptomycin did not.5 After the experimental study reported by47 all clinically affected or seropositive herds in the same region in Namibia were treated with fluoroquinolone and monitored clinically for two years; an immediate drop in cases was observed in the first three months after treatment, with no further cases reported during the two-year period.77
Guidelines on antimicrobial treatment of cases of CBPP are therefore required before that approach can be formally recommended as a control measure, possibly in combination with the other measures, such as vaccination and slaughter.36 It is likely, however, that antibiotic therapy will continue to be used to treat animals with respiratory signs because antibiotics are very efficiently distributed by the private sector to livestock owners, and there are reports of misuse of these products, including in treatment of CBPP.2, 3, 51 However, there is a lack of qualitative and quantitative information on the use of antibiotics in Africa. Bans on the use of antimicrobial therapy are unlikely to be respected in many locations, which poses an additional risk for the emergence of antibiotic resistance. Such resistance emergence has been witnessed in Europe for another mycoplasma affecting cattle, M. bovis.39 Nevertheless, strategic and targeted use of antibiotic therapy as a tool in integrated programmes for the eradication of CBPP deserves further investigation.61, 77
Antibiotic therapy should, however, be used in the case of post-vaccinal reactions at the site of the inoculation that develop in a small number of cases and could jeopardize the faith of cattle owners in the efficacy of vaccination.
Vaccination is very important in the control of CBPP in endemic areas, especially in countries where animal movement control is impractical. Attenuated vaccines against CBPP have been developed by in vitro culture of the organism or culture in unnatural hosts such as embryonated hens’ eggs.93
Attenuated virulent strains stimulate immunity best63 but they also induce the most severe and undesirable local and systemic reactions refer to as "Willems reaction” (Figure 18) that may even result in the death of the animal.86 A compromise between virulence, immunogenicity and safety has been achieved In the attenuated vaccines currently in use.15
Until the 1950s it was commonly thought that if vaccination were to be effective, a local lesion needed to be induced at the site of inoculation. This situation changed at the insistence of cattle owners because of the severe post-vaccination reactions (Figure 18) and mortality in their cattle. The 44th egg-passage of strain T1, named T1/44, is sufficiently attenuated to protect cattle without severe post-vaccinal reactions at the injection site; however, such reactions may still rarely occur in the field.
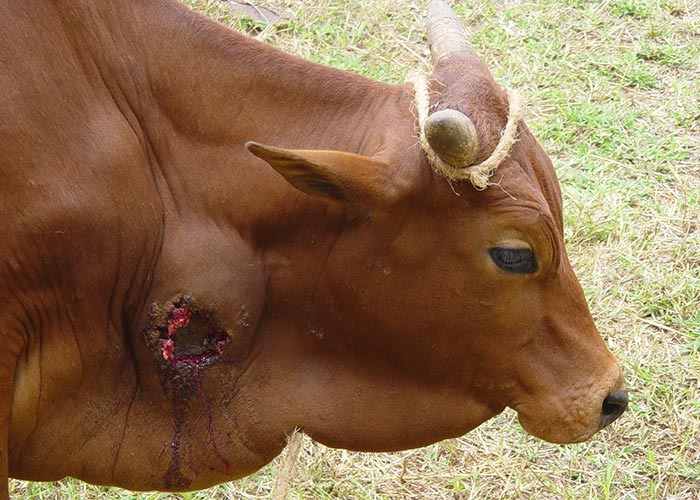
Figure 18 Severe necrotizing and ulcerative skin lesion refer to as "Willems reaction" following subcutaneous vaccination with T1/44 strain. Note severe oedema of the dewlap. (By courtesy of F. Thiaucourt and H Wesonga (KARI), Campus de Baillarguet, ASTRE, TA A117, Cedex 5 Montpellier, France, 34398)
Among the many vaccine strains that have been developed only two are now used in Africa, T1/44 and its streptomycin-resistant derivate, T1sr.83 Vaccination failures occurring in East and southern Africa in 1994 cast doubt on the efficacy of the T1sr strain although it was identified as the preferred choice by a panel of experts in 1990.
The protection rate afforded by a vaccine strain is difficult to establish experimentally for several reasons. Firstly, experimental reproduction of CBPP is difficult and infection by intratracheal intubation may result in only few animals being infected. Transmission experiments preferably need to be performed by contact between infected and uninfected animals. Therefore, large numbers of cattle must be used and the reproducibility of the rate of transmission from one experiment to another is difficult to achieve. It is even more difficult to evaluate the protection afforded by a vaccine by observations in the field, as a number of factors potentially interfere with the efficacy of the vaccine, e.g. improper storage of the freeze-dried product, use of unsuitable diluents, and reconstituted vaccine not being inoculated within a reasonable time.
The question of identity of the T1sr strain has been resolved by the use of molecular tools that enabled its characterization and particularly its distinction from the less protective KH3J strain.58, 100 In 1998 and 1999 the two strains, T1/44 and T1sr, were thoroughly re-evaluated.102, 116 The experimental protocols used included vaccination of naive cattle at the minimum dose (107 live mycoplasmas per dose) recommended by the OIE. The trial was conducted in three different locations in Africa. Animals were challenged with virulent organisms three months after vaccination. Results confirmed and re-established the existing knowledge on vaccine efficacy: primary vaccination induced only 40 to 60 per cent protection, whichever vaccine strain was used. The duration of immunity was longer with T1/44 than with T1sr, and revaccination after one year enhanced the level of protection to 80 to 95 per cent. It has therefore been re-established that satisfactory protection by vaccination can be achieved only by repeated vaccination, and those concerned with control strategies based on vaccination need to take that into account. The protection rate afforded by vaccines was similar in all the locations, indicating that genomic variation is not associated with pathogenic strains evading the immune reaction following vaccination with T1/44.
The vaccine strain T1sr is completely avirulent, but the T1/44 strain may induce a post-vaccinal reaction at the injection site in some animals. The number of reactors is unpredictable and the introduction of T1/44 in vaccination strategies has to be ‘played by ear’, which means that vaccinating teams must be prepared to treat the reactors with antibiotics, if required, two to three weeks after vaccination. The number of reactors tends to be smaller following re-vaccination. Whichever strain is used, the seroconversion rate measured by CFT or cELISA is very variable, and there is no strict correlation between the test results and the protection obtained. Animals that do seroconvert should be protected from developing the disease but those that do not seroconvert may also be protected. There is an urgent need for a test that allows the level of protection to be measured, as it would permit the efficacy of the vaccination campaigns to be managed. However, such a tool, which was an important component for the control of rinderpest, is not available for CBPP.
The basis of protection in CBPP is not well understood, and neither is the identity of Mmm antigens that elicit protection. Furthermore, the nature of the immune response that results in protection is also unclear.46 CD4 T-cells secreting gamma interferon may play an important role in the latter respect.
Control strategies require strong support from governments and active participation on the part of cattle owners. Eradication strategies also require the full co-ordination of neighbouring states, as CBPP is a transboundary disease. A slaughter-out policy was successfully applied in Botswana from 1995 to 1997 to eradicate CBPP. This required not only a considerable investment for compensating the owners of slaughtered animals and for restocking purposes, but it also required strict control of animal movements to prevent reintroduction of the disease.
Annual blanket vaccination certainly results in a dramatic reduction of CBPP outbreaks, but vaccination may fail to eradicate the disease if there is no way to control the reintroduction of infected cattle from neighbouring regions or countries. This was the situation prevailing in northern Namibia in 2001, and combined vaccination campaigns in that country and southern Angola are the only way to solve the problem. In Australia, blanket vaccination was used to reduce CBPP prevalence before regular serological testing and slaughter procedures were available; these eventually led to the eradication of CBPP in 1973.44 Vaccination, combined with heightened surveillance, resulted in the eradication of CBPP from China by 1989 after the programme was initiated in 1956 with production of the first vaccine.115
Treatment with antimicrobials might also be seen as an additional tool for the control of CBPP, but lack of information on its long-term impact is a constraint. However, as indicated above, there is evidence that inclusion of appropriate therapy in control and eradication programmes could prove beneficial.22, 61, 77 Cost-effective control strategies should include the combined use of available measures, the choice of which will depend on the local situation.
It is of utmost importance for countries free of CBPP, but under risk of importing the disease, to maintain diagnostic capability for early detection and surveillance.
References
- ABDO, E.M., NICOLET, J. & FREY J., 2000. Antigenic and genetic characterization of lipoprotein LppQ from Mycoplasma mycoides subsp. mycoides. Clinical and Diagnostic Laboratory Immunology, 7, 588–595.
- ADESOKAN, H.K., AGADA, C.A., ADETUNJI, V.O. & AKANBI, I.M., 2013. Oxytetracycline and penicillin-G residues in cattle slaughtered in south-western Nigeria: Implications for livestock disease management and public health. Journal of the South African Veterinary Association, 84(1), Art. #945, 5 pages. (http://dx.doi.org/10.4102/jsava.v84i1.945).
- ADESOKAN, H.K., AKANBI, I.O., AKANBI, I.M. & OBAWEDA, R.A., 2015. Patterns of antimicrobial usage in livestock animals in south-western Nigeria: The need for alternative plans. Onderstepoort Journal of Veterinary Research, 82(1), Art. #816, 6 pages. (http://dox.doi.org/10.4102/ojvr.v82i1.816).
- AKAFEKWA, G.I., 1975. The diagnosis, control and eradication of contagious bovine pleuropneumonia in Zambia. Bulletin of Epizootic Diseases of Africa, 84, 429–449.
- ALMAW, G., DUGUMA, M., WUBETIE, A., TULI, G. & KORAN, T., 2016. A contagious bovine pleuropneumonia outbreak on a research farm in Ethiopia, and its dynamics over an eight-month period. Revue Scientifique et Technique de Office international des Épizooties, 35(3), 787-793.
- AMANFU, W., 2006. The use of antibiotics for CBPP control: the challenges. In: LUBROTH, J., (ed) CBPP control: antibiotics to the rescue, vol 8. Animal production and health proceedings. FAO, Rome, 7-11
- AMANFU, W., SEDIADIE, S., MASUPU, K.V., BENKIRANE, A., GEIGER, R. & THIAUCOURT, F., 1998. Field validation of a competitive ELISA for the detection of contagious bovine pleuropneumonia in Botswana. Revue D'Elevage et De Medecine Veterinaire Des Pays Tropicaux, 51, 189–193
- AMANFU, W., 2009. Contagious bovine pleuropneumonia (lungsickness) in Africa. Onderstepoort Journal of Veterinary Research, 76(1), 13-17.
- AMANFU, W., MASUPU, K.V., ADOM, E.K., RABOROKGWE, M.V. & BASHIRUDDIN, J., 1998. An outbreak of contagious bovine pleuropneumonia in Ngamiland district of north-western Botswana. The Veterinary Record, 143, 46–48.
- ANDERSON, R.M. & MAY, R.M., 1982. Directly transmitted infectious diseases: Control by vaccination. Science, 215, 1053-1060.
- ANON., 1938–1987. Annual Reports of the Division of Veterinary Services, Private Bag 12022, Windhoek, Namibia.
- ARFI, Y., MINDER, L., DI PRIMO, C., LE ROY, A., EBEL, C., COQUET, L., CLAVEROL, S., VASHEE, S., JORES, J., BLANCHARD, A. & SIRAND PUGNET, P., 2016. MIB–MIP is a mycoplasma system that captures and cleaves immunoglobulin G. Proceedings of the National Academy of Sciences, 113, 5406-5411.
- AYE, R., MWIRIGI, M.K., FREY, J., PILO, P., JORES, J. & NAESSENS, J., 2015. Cyto-adherence of Mycoplasma mycoides subsp. mycoides to bovine lung epithelial cells. BMC Veterinary Research, 11, 27.
- AYLING, R.D., BAKER, S.E., NICHOLAS, R.A.J., PEEK, M.L. & SIMON, A.J., 2000. Comparison of in vitro activity of danofloxacin, florfenicol, oxytetracyclin, spectinomycin and tilmicosin against Mycoplasma mycoides subsp. mycoides small colony type. The Veterinary Record, 146, 243–246.
- BARILE, M.F., 1985. Immunization against mycoplasma infections. In: RAZIN, S. & BARILE, M.F., (eds). The Mycoplasmas. Vol. IV. Orlando, Florida: Academic Press Inc.
- BASHIRUDDIN, J.B., NICHOLAS, R.A.J., SANTINI, F.G., READY, R.A., WOODWARD, M.J. & TAYLOR, T.K., 1994. Use of the polymerase chain reaction to detect Mycoplasma DNA in cattle with contagious pleuropneumonia. Veterinary Record, 134, 240–241.
- BASHIRUDDIN, J.B., DE SANTIS, P., VACCIANA, A. & SANTINI, F.G., 1999. Detection of Mycoplasma mycoides subspecies mycoides SC in clinical material by a rapid colorimetric PCR. Molecular and Cellular Probes, 13, 23–28.
- BASHIRUDDIN, J.B., SANTINI, F.G., DE SANTIS, P., VISAGGIO, M.C., DI FRANCESCO, G., D’ANGELO, A. & NICHOLAS, R.A.J., 1999. Detection of Mycoplasma mycoides subspecies mycoides in tissues from an outbreak of contagious bovine pleuropneumonia by culture, immunohistochemistry and polymerase chain reaction. Veterinary Record, 145, 271–274.
- BASHIRUDDIN, J.B., TAYLOR, T.K. & GOULD, A.R., 1994. A PCR-based test for the specific identification of Mycoplasma mycoides subspecies mycoides SC. Journal of Veterinary Diagnostic and Investigation, 6, 28–434.
- BELLINI, S., GIOVANNINI, A., DI FRANCESCO, C., TITTARELLI, M. & CAPORALE, V., 1998. Sensitivity and specificity of serological and bacteriological tests for contagious bovine pleuropneumonia. Revue Scientifique et Technique de Office international des Épizooties, 17(3), 654-659.
- BERTIN, C., PAU-ROBLOT, C., COURTOIS, J., MANSO-SILVÁN, L., TARDY, F., POUMARAT, F., CITTI, C., SIRAND-PUGNET, P., GAURIVAUD, P. & THIAUCOURT, F., 2015. Highly dynamic genomic loci drive the synthesis of two types of capsular or secreted polysaccharides within the Mycoplasma mycoides cluster. Applied and Environmental Microbiology, 81, 676-687.
- BRUNNER, H. & LABER, G., 1985. Chemotherapy of mycoplasma infections. In: RAZIN, S. & BARILE, M.F., (eds). The Mycoplasmas. Vol. IV. Orlando, Florida: Academic Press Inc.
- BYGRAVE, A.C., MOULTON, J.E. & SHIFRINE, M., 1968. Clinical, serological and pathological findings in an outbreak of contagious bovine pleuropneumonia. Bulletin of Epizootic Diseases of Africa, 16, 21–46.
- CAMPBELL, A.D. & TURNER A.W., 1953. Studies on contagious bovine pleuropneumonia of cattle. IV. An improved complement fixation test. Australian Veterinary Journal, 29, 154–163.
- CHENG, X., NICOLET, J., POUMARAT, F., REGALLA, J., THIAUCOURT, F. & FREY, J., 1995. Insertion element IS1296 in Mycoplasma mycoides subsp. mycoides small colony identifies a European clonal line distinct from African and Australian strains. Microbiology, 141, 3221–3228.
- CHIMA, J.C. & PAM, G., 1985. Contagious bovine pleuropneumonia: A comparison between the passive haemagglutination test and the complement fixation test. Revue Scientifique et Technique de Office international des Épizooties, 4, 517–522.
- CITTI, C. & ROSENGARTEN, R., 1997. Mycoplasma genetic variation and its implication for pathogenesis. Wiener Klinische Wochenschrift, 109/14– 15, 562–568.
- CLAY, A.L. & LLOYD, L.C., 1975. The eradication of contagious bovine pleuropneumonia from Australia. Bulletin of the Office international des Épizooties, 84, 463-475.
- COETZER, J.A.W. & PICARD, J.A., 2001. Faculty of Veterinary Science, University of Pretoria. Personal communication.
- COTTEW, G.S., BREARD, A., DAMASSA, A.J., ERNØ, H., LEACH, R.H., LEFEVRE, P.C., RODWELL, A.W. & SMITH, G.R., 1987. Taxonomy of the Mycoplasma mycoides cluster. Israel Journal of Medical Science, 23, 632–635.
- CURASSON, G., 1942. Péripneumonie bovine. In: Traité de Pathologie exotique Vétérinaire et Compareé, 2, 276–353. 2nd Edition. Paris VI: Vigot Frères Ed.
- DAVIES, G. & GILBERT, F.R., 1969. Contagious bovine pleuropneumonia vaccination in East Africa. Bulletin of Epizootic Diseases of Africa, 17, 21–26.
- DEDIEU, L., MADY, V. & LEFEVRE, P.C., 1994. Development of a selective polymerase chain reaction assay for the detection of Mycoplasma mycoides subsp. mycoides SC (contagious bovine pleuropneumonia agent). Veterinary Microbiology, 42, 327–339.
- DUPUY, V., MANSO-SILVÁN, L., BARBE, V., THEBAULT, P., DORDET-FRISONI, E., CITTI, C., POUMARAT, F., BLANCHARD, A., BRETON, M., SIRAND-PUGNET, P. & THIAUCOURT, F., 2012. Evolutionary history of contagious bovine pleuropneumonia using next generation sequencing of Mycoplasma mycoides subsp mycoides "Small Colony". PLoS ONE, 7, e46821.
- EGWU, G.O., NICHOLAS, R.A.J., AMEH, J. A. & BASHIRUDDIN, J.B., 1996. Contagious bovine pleuropneumonia: an update. Veterinary Bulletin, 66, 875–888.
- FAO., 2007. CBPP control: antibiotics to the rescue? FAO-OIE-AU/IBAR-IAEA Consultative Group Meeting on CBPP in Africa, Rome, 6-8 November 2006. FAO Animal Health and Production Proceedings No. 8, Food and Africulture Organization of the United Nations, Rome.
- GABRIDGE, M.G., CHANDLER, D.K.F. & DANIELS, M.J., 1985. Pathogenicity factors in mycoplasmas and spiroplasmas. In: RAZIN, S. & BARILE, M.F., (eds). The Mycoplasmas, Vol. IV. Orlando, Florida: Academic Press Inc.
- GAURIVAUD, P., LAKHDAR, L., LE GRAND, D., POUMARAT, F. & TARDY, F., 2014. Comparison of in vivo and in vitro properties of capsulated and noncapsulated variants of Mycoplasma mycoides subsp. mycoides strain Afadé: a potential new insight into the biology of contagious bovine pleuropneumonia. FEMS Microbiology Letters, 359, 42-49.
- GAUTIER-BOUCHARDON, A.V., FERRÉ, S., LE GRAND, D., PAOLI, A., GAY, E. & POUMARAT F., 2014. Overall decrease in the susceptibility of Mycoplasma bovis to antimicrobials over the past 30 years in France. PLoS ONE, 9(2), e87672.
- GIOVANNINI, A., BELLINI, S., SALMAN, M.D. & CAPORALE, V., 2000. Spatial risk factors related to outbreaks of contagious bovine pleuropneumonia in northern Italy (1990-1993). Revue Scientifique et Technique, Office international des Épizooties, 19, 764-772.
- GONÇALVES, R., 1994. Isolation and identification of Mycoplasma mycoides subsp. mycoides SC from bull semen and sheath washings in Portugal. The Veterinary Record, 135, 308–309.
- GONÇALVES, R., REGALLA, J, NICOLET J, FREY J, NICHOLAS R, BASHIRUDDIN J, DE SANTIS P. & GONCALVES, A.P., 1998. Antigen heterogeneity among Mycoplasma mycoides subsp. mycoides SC isolates: discrimination of major surface proteins. Veterinary Microbiology, 63, 13–28.
- GRIFFIN, R.M., 1965. A gel diffusion precipitin test for contagious bovine pleuropneumonia. Journal of Comparative Pathology, 75, 223–231.
- HALLER, A., 1773. Mémoire sur la contagion parmi le bétail mis au jour pour l'instruction du public (Berne), 32.
- HENNING, M.W., 1956. Animal Diseases in South Africa. 3rd Edition. Pretoria: Central News Agency Ltd.
- HOWARD, C.J. & TAYLOR, G., 1985. Humoral and cell-mediated immunity. In: RAZIN, S. & BARILE, M.F., (eds). The Mycoplasmas, Vol. IV. Orlando, Florida: Academic Press Inc.
- HÜBSCHLE, O.J.B., AYLING, R.D., GODINHO, K., LUKHELE, O., TJIPURA-ZAIRE, G., ROWAN, T.G. & NICHOLAS, R.A.J., 2006. Danofloxacin (AdvocinTM) reduces the spread of contagious bovine pleuropneumonia to healthy in-contact cattle. Research in Veterinary Science, 81, 304-309.
- HUDSON, J.R., 1968. Contagious bovine pleuropneumonia. Experiments on the susceptibility and protection by vaccination of different types of cattle. Australian Veterinary Journal, 44(3), 83-89.
- HUDSON, J.R., 1971. Contagious bovine pleuropneumonia. Food and Agricultural Organization Agricultural Studies, No. 86. Rome: Food and Agricultural Organization of the United Nations.
- HUDSON, R. & TURNER, A.W., 1963. Contagious bovine pleuropneumonia: a comparison of the efficacy of two types of vaccine. Australian Veterinary Journal, 39, 373-385.
- KAIRU-WANYOIKE, S.W., KIARA, H., HEFFERNAN, C., KAITIBIE, S., GITAU, G.K., MCKEEVER, D. & TAYLOR, N.M., 2014. Control of contagious bovine pleuropneumonia: Knowledge, attitudes, perceptions and practices in Narok district of Kenya. Preventive Veterinary Medicine, 115, 143-156.
- KUSILUKA, L.J., OJENIYI, B., FRIIS, N.F., KAZWALA, R.R. & KOKOTOVIC, B., 2000. Mycoplasmas isolated from the respiratory tract of cattle and goats in Tanzania. Acta Veterinaria Scandinavica, 41, 299–309.
- LE GOFF, C. & THIAUCOURT, F., 1998. A competitive ELISA for the specific diagnosis of contagious bovine pleuropneumonia (CBPP). Veterinary Microbiology, 60, 179–191.
- LEACH, R.H., ERNØ, H. & MACOWAN, K.J., 1993. Proposal for the designation of F38-type caprine mycoplasmas as Mycoplasma capricolum subsp. capripneumoniae subsp. nov. and consequent obligatory relegation of strains currently classified as M. capricolum to an additional subspecies, M. capricolum subsp. capricolum subsp. nov. International Journal of Systematic Bacteriology, 43, 603–605.
- LEACH, T.M., 1957. The occurrence of contagious bovine pleuropneumonia in species other than domesticated cattle. Bulletin of Epizootic Diseases of Africa, 5, 325–328.
- LESNOFF, M., LAVAL, G., BONNET, P., CHALVET-MONFRAY, K., LANCELOT, R. & THIAUCOURT, F., 2004. A mathematical model of the effects of chronic carriers on the within-herd spread of contagious bovine pleuropneumonia in an African mixed crop-livestock system. Preventive Veterinary Medicine, 62, 101-117.
- LI, Y., WANG, Y., WANG, R., ZHU, Y., LIU, S., WANG, Q., SHAO, J., CHEN, Y., GAO, L., ZHOU, C., LIU, H., WANG, X., ZHENG, H. & XIN, J., 2016. Changes in pathogenicity and immunogenicity of Mycoplasma mycoides subsp. mycoides strains revealed by comparative genomics analysis. Scientific Reports, 6, 19081.
- LORENZON, S., DAVID, A., NADEW, M., WESONGA, H. & THIAUCOURT, F., 2000. Specific PCR identification of the T1 vaccine strains for contagious bovine pleuropneumonia. Molecular and Cellular Probes, 14, 205–210.
- MANSO-SILVÁN, L., VILEI, E.M., SACHSE, K., DJORDJEVIC, S.P., THIAUCOURT, F. & FREY, J., 2009. Proposal to assign Mycoplasma leachii sp nov as a new species designation for Mycoplasma sp bovine group 7 of Leach and to reclassify Mycoplasma mycoides subsp mycoides LC as a serovar of Mycoplasma mycoides subsp. capri. International Journal of Systematic and Evolutionary Microbiology, 59, 1353-1358.
- MARCH, J.B., CLARK, J. & BRODLIE, M., 2000. Characterization of strains of Mycoplasma mycoides subsp mycoides small colony type isolated from recent outbreaks of contagious bovine pleuropneumonia in Botswana and Tanzania: Evidence for a new biotype. Journal of Clinical Microbiology, 38, 1419–1425.
- MARINER, J.C., MCDERMOTT, J., HEESTERBEEK, J.A.P., THOMSON, G. & MARTIN, S.W., 2006. A model of contagious bovine pleuropneumonia transmission dynamics in East Africa. Preventive Veterinary Medicine, 73, 55-74.
- MASIGA, W.N. & READ, W.C.S., 1972. Comparative susceptibility of Bos indicus and Bos taurus to contagious bovine pleuropneumonia and the efficacy of the T1 broth culture vaccine. The Veterinary Record, 90, 499–502.
- MASIGA, W.N. & WINDSOR, R.S., 1975. Immunity to contagious bovine pleuropneumonia. The Veterinary Record, 97, 350–351.
- MASIGA, W.N. & WINDSOR, R.S., 1978. Some evidence of an age susceptibility to contagious bovine pleuropneumonia. Research in Veterinary Science, 24, 328–333.
- MASIGA, W.N., DOMENECH, J. & WINDSOR, R.S., 1996. Manifestation and epidemiology of contagious bovine pleuropneumonia in Africa. Revue Scientifique et Technique de Office international des Épizooties, 15, 1283–1308.
- MASIGA, W.N., WINDSOR, R.S. & READ, W.C.S., 1972. A new mode of spread of contagious bovine pleuropneumonia. The Veterinary Record, 90, 247–248.
- MBENGUE, M., DIALLO, A.A., LO, F.T., LO, M.M., DIOP, M., SECK, P.S., SAMB, Y., DIOUF, M. & THIONGANE, Y., 2013. Réémergence de la péripneumonie contagieuse bovine au Sénégal. Bulletin de la Société de Pathologie Exotique, 106, 212-215.
- MCINTYRE, G., 1939. The lungsickness campaign in the Caprivi-Zipfel. Journal of the South African Veterinary Medical Association, 10, 4–12.
- MENDES, A.M., 1953. Contagious bovine pleuropneumonia control in Angola. Bulletin of Epizootic Diseases of Africa, 1, 426–432.
- MISEREZ, R., PILLOUD, T., CHENG, X., NICOLET, J., GRIOT, C. & FREY, J., 1997. Development of a sensitive nested PCR method for the specific detection of Mycoplasma mycoides subsp. mycoides SC. Molecular and Cellular Probes, 11, 103–111.
- MUUKA, G., HANG'OMBE, B.M., NALUBAMBA, K.S., KABILIKA, S., MWAMBAZI, L. & MUMA, J.B., 2011. Comparison of complement fixation test competitive ELISA and LppQ ELISA with post-mortem findings in the diagnosis of contagious bovine pleuropneumonia (CBPP). Tropical Animal Health and Production, 43(5), 1057-1062.
- NEWING, C.R., 1955. An improved method for the preparation of an antigen for the rapid slide test for contagious bovine pleuro-pneumonia. British Veterinary Journal, 111, 378–384.
- NEWTON, L.G., 1992. Contagious bovine pleuropneumonia in Australia: Some historic highlights from entry to eradication. Australian Veterinary Journal, 69, 306–317.
- NIANG, M., DIALLO, M., CISSE, O., KONE, M., DOUCOURE, M., ROTH, J.A., BALCER-RODRIGUES, V. & DEDIEU, L., 2006. Pulmonary and serum antibody responses elicited in zebu cattle experimentally infected with Mycoplasma mycoides subsp. mycoides SC by contact exposure. Veterinary Research, 37(5), 733-744.
- NIANG, M., SERY, A., DOUCOURÉ, M., KONÉ, M., N'DIAYE, M., AMANFU, W. & THIAUCOURT, F., 2010. Experimental studies on long-acting tetracycline treatment in the development of sequestra in contagious bovine pleuropneumonia - infected cattle. Journal of Veterinary Medicine and Animal Health, 2(4), 35-45.
- NICHOLAS, R., BASHIRUDDIN, J., AYLING, R. & MILES, R., 2000. Contagious bovine pleuropneumonia: a review of recent developments. Veterinary Bulletin, 70, 827–838.
- NICHOLAS, R.A.J., AYLING, R.D., TJIPURA-ZAIRE, G. & ROWAN, T., 2012. Treatment of contagious bovine pleuropneumonia. Veterinary Record, 171, 510-511.
- OTAKE, S., DEE, S., CORZO, C., OLIVEIRA, S. & DEEN, J., 2010. Long-distance airborne transport of infectious PRRSV and Mycoplasma hyopneumoniae from a swine population infected with multiple viral variants. Veterinary Microbiology, 145, 198-208.
- PEIRES, J.B., 1989. The Dead will arise. Johannesburg: Ravan Press.
- PERSSON, A., PETTERSSON, B., BÖLSKE, G. & JOHANSSON, K.E., 1999. Diagnosis of contagious bovine pleuropneumonia by pcr-laser-induced fluorescence and pcr-restriction endonuclease analysis based on the 16s rrna genes of mycoplasma mycoides subsp mycoides SC. Journal of Clinical Microbiology, 37, 3815–3821.
- PILO, P., FREY, J. & VILEI, E.M., 2007. Molecular mechanisms of pathogenicity of Mycoplasma mycoides subsp mycoides SC. Veterinary Journal, 174, 513-521.
- POUMARAT, F. & SOLSONA, M., 1995. Molecular epidemiology of Mycoplasma mycoides subsp. mycoides biotype Small Colony, the agent of contagious bovine pleuropneumonia. Veterinary Microbiology, 47, 305–315.
- PROVOST, A., 1982. Advantages of a joint campaign against rinderpest and bovine pleuropneumonia. Revue Scientifique et Technique de Office international des Épizooties, 1, 687–704.
- PROVOST, A., PERREAU, P., BREARD, A., LE GOFF, C., MARTEL, J.L. & COTTEW, G.S., 1987. Contagious bovine pleuropneumonia. Revue Scientifique et Technique de Office international des Épizooties, 6, 625–679.
- REGALLA, J., CAPORALE, V., GIOVANNINI, A., SANTINI, F., MARTEL, J.L. & GONÇALVES, A.P., 1996. Manifestation and epidemiology of contagious bovine pleuropneumonia in Europe. Revue Scientifique et Technique de Office international des Épizooties, 15, 1309–1329.
- REVELL, S.G., 1973. Local reactions following CBPP vaccination in Zambia. Tropical Animal Health and Production, 5, 246–252.
- RURANGIRWA, F.R., SHOMPOLE, P.S., WAMBUGU, A.N. & MCGUIRE, T.C., 2000. Monoclonal antibody differentiation of Mycoplasma mycoides subsp. mycoides small-colony strains causing contagious bovine pleuropneumonia from less important large-colony strains. Clinical and Diagnostic Laboratory Immunology, 7(3), 519-521.
- SANTINI, F.G., D’ANGELO, A.R., SCACCHIA, M., VISAGGIO, M., FARINELLI, G., DI FRANSCESCO, G. & GUARDUCCI, M., 1992. Pulmonary sequestrum from Mycoplasma mycoides var. mycoides SC in a domestic buffalo: Isolation, anatomo-histology and immunohistochemistry. Veterinaria Italiana, 4, 4–10.
- SCANZIANI, E., PALTRINIERI, S., BOLDINI, M., GRIECO, V., MONACI, C., GIUSTI, A.M. & MANDELLI, G., 1997. Histological and immunohistochemical findings in thoracic lymph nodes of cattle with contagious bovine pleuropneumonia. Journal of Comparative Pathology, 117, 127–136.
- SCHIECK, E., LILJANDER, A., HAMSTEN, C., GICHERU, N., SCACCHIA, M., SACCHINI, F., HELLER, M., SCHNEE, C., STERNER-KOCK, A., HLINAK, A., NAESSENS, J., POOLE, J., PERSSON, A. & JORES, J., 2014. High antibody titres against predicted Mycoplasma surface proteins do not prevent sequestration in infected lung tissue in the course of experimental contagious bovine pleuropneumonia. Veterinary Microbiology, 172, 285-293.
- SCHNEIDER, H.P., 1987. 100 years of veterinary legislation and early forms of disease control in South West Africa/Namibia. Proceedings of the XXIII World Veterinary Congress, Montreal, Canada.
- SHAW, C.D., 1971. Zambia. Contagious bovine pleuropneumonia 1970 outbreak. Progress report. Bulletin de l’Office International des Epizooties, 76, 673–677.
- SHERIFF, D. & PIERCY, S.E., 1952. Experiments with avianised strain of the organism of contagious bovine pleuropneumonia. The Veterinary Record, 64, 615–621.
- SHIFRINE, M., STONE, S.S. & STAAK, C., 1970. Contagious bovine pleuropneumonia in African buffalo (Syncerus caffer). Bulletin of Epizootic Diseases of Africa, 18, 201–205.
- SRIVASTAVA, N.C., THIAUCOURT, F., SINGH, V.P., SUNDER, J. & SINGH, V.P., 2000. Isolation of Mycoplasma mycoides small colony type from contagious caprine pleuropneumonia in India. The Veterinary Record, 147, 520–521.
- STÄRK, K.D.C., VICARI, A., KIHM, U. & NICOLET, J., 1995. Surveillance of contagious bovine pleuropneumonia in Switzerland. Revue Scientifique et Technique de Office international des Épizooties, 14, 621–629.
- STERNER-KOCK, A., HAIDER, W., SACCHINI, F., LILJANDER, A., MEENS, J., POOLE, J., GUSCHLBAUER, M., HELLER, M., NAESSENS, J. & JORES, J., 2016. Morphological characterization and histochemical detection of proinflammatory cytokines IL-1β, IL-17A and TNF-α in lung lesions associated with contagious bovine pleuropneumonia. Tropical Animal Health and Production, 48, 569-576.
- STRADAIOLI, G., SYLLA, L., MAZZARELLI, F., ZELLI, R., RAWADI, G. & MONACI, M., 1999. Mycoplasma mycoides subsp. mycoides SC identification by PCR in sperm of seminal vesiculitis-affected bulls. Veterinary Research, 30, 457–466.
- THIAUCOURT, F., LORENZON, S., DAVID, A. & BREARD, A., 2000. Phylogeny of the Mycoplasma mycoides cluster as shown by sequencing of a putative membrane protein gene. Veterinary Microbiology, 72, 251–268.
- THIAUCOURT, F., LORENZON, S., DAVID, A., TULASNE, J.J. & DOMENECH, J., 1998. Vaccination against contagious bovine pleuropneumonia and the use of molecular tools in epidemiology. Annals of the New York Academy of Science, 849, 146–151.
- THIAUCOURT, F., MANSO-SILVÁN, L., WOUBIT, S., BARBE, V., VACHERIE, B., JACOB, D., BRETON, M., DUPUY, V., LOMENECH, A.M., BLANCHARD, A. & SIRAND-PUGNE, T.P., 2011. Mycoplasma mycoides from “mycoides Small Colony” to “capri” A micro evolutionary perspective. BMC Genomics, 12, 114.
- THIAUCOURT, F., YAYA, A., WESONGA, H., HUEBSCHLE, O.J.B., TULASNE, J.J. & PROVOST, A., 2000. Contagious bovine pleuropneumonia: A reassessment of the efficacy of vaccines used in Africa. Annals of the New York Academy of Science, 916, 71–80.
- THOMSON, G.R. & PENRITH, M.L., 2017. Eradication of transboundary animal diseases: Can the rinderpest story be repeated? Transboundary and Emerging Diseases, 64, 459-475.
- TOTTÉ, P., PUECH, C., RODRIGUES, V., BERTIN, C., MANSO-SILVÁN, L. & THIAUCOURT, F., 2015. Free exopolysaccharide from Mycoplasma mycoides subsp. mycoides possesses anti-inflammatory properties. Veterinary Research, 46, 122.
- TRETHEWIE, E.R. & TURNER, A.W., 1961. Preventive tail-tip inoculation of calves against bovine contagious pleuropneumonia. II. Vegetative endocarditis (valvitis) and myocarditis as sequelae to post-inoculation arthritis. Australian Veterinary Journal, 37, 27–36.
- TURNER, A.W. & TRETHEWIE, E.R., 1961. Preventive tail-tip inoculation of calves against bovine contagious pleuropneumonia. I. Influence of age upon inoculation upon tail reactions, serological responses, and the incidence of swollen joints. Australian Veterinary Journal, 37, 1–8.
- VILEI, E.M. & FREY, J., 2001. Genetic and biochemical characterization of glycerol uptake in Mycoplasma mycoides subsp. mycoides SC: Its impact on H2O2 production and virulence. Clinical and Diagnostic Laboratory Immunology, 8, 85–92.
- VILEI, E.M., ABDO, E.M., NICOLET, J., BOTELHO, A., GONÇALVES, R. & FREY, J., 2000. Genomic and antigenic differences between the European and African/Australian clusters of Mycoplasma mycoides subsp. mycoides SC. Microbiology, 146, 477–486.
- VILEI, E.M., NICOLET, J. & FREY, J., 1999. IS1634, a novel insertion element creating long, variable-length direct repeats which is specific for Mycoplasma mycoides subsp. mycoides small-colony type. Journal of Bacteriology, 181, 1319–1323.
- WEISBURG, W.G., TULLY, J.G., ROSE, D.L., PETZEL, J.P., OYAIZU, H., YANG, D., MANDELCO, L., SECHREST, J., LAWRENCE, T.G., VAN ETTEN J., MANILOFF, J. & WOESE C.R. 1989. A phylogenetic analysis of the mycoplasmas: Basis for their classification. Journal of Bacteriology, 171, 6455–6467.
- WESTBERG, J., PERSSON, A., HOLMBERG, A., GOESMANN, A., LUNDEBERG, J., JOHANSSON, K.E., PETTERSSON, B. & UHLEN, M., 2004. The genome sequence of Mycoplasma mycoides subsp. mycoides SC type strain PG1T the causative agent of contagious bovine pleuropneumonia (CBPP). Genome Research, 14, 221-227.
- WILLEMS, L., 1852. Mémoire sur la pleuro-pneumonie Épizootique du gros bétail. Recueil de Médecine Vétérinaire Pratique, 9, 401–434.
- WINDSOR, R.S. & MASIGA, W.N., 1977. Investigations into the role of carrier animals in the spread of contagious bovine pleuropneumonia. Research in Veterinary Science, 23, 224–229.
- WISE, K.S., CALCUTT, M.J., FOECKING, M.F., MADUPU, R., DEBOY, R.T., RÖSKE, K., HVINDEN, M.L., MARTIN, T.R., DURKIN, A.S., GLASS, J.I. & METHÉ, B.A., 2012. Complete genome sequences of Mycoplasma leachii strain PG50(T) and the pathogenic Mycoplasma mycoides subsp. mycoides Small Colony biotype strain Gladysdale. Journal of Bacteriology, 194, 4448-4449.
- XIN, J., LI, Y., NICHOLAS, R.A.J., CHEN, C., LIU, Y., ZHANG, M. & DONG, H., 2012. A history of the prevalence and control of contagious bovine pleuropneumonia in China. Veterinary Journal, 191, 166-170.
- YAYA, A., GOLSIA, R., HAMADOU, B., AMARO, A. & THIAUCOURT, F., 1999. Essai comparatif d’efficacité des deux souches vaccinales T1/44 et T1sr contre la péripneumonie contagieuse bovine. Revue d’Élevage Neet de Médecine Vétérinaire des Pays Tropicaux, 52, 171–179.
- YAYA, A., MANSO-SILVÁN, L., BLANCHARD, A. & THIAUCOURT, F., 2008. Genotyping of Mycoplasma mycoides subsp mycoides SC by multilocus sequence analysis allows molecular epidemiology of contagious bovine pleuropneumonia. Veterinary Research, 39, 14.
- YAYA, A., HAMADOU, B., YAYA, D., ABDOULKADIRI, S. & THIAUCOURT, F., 2000. Inoculation expérimentale de l'agent de la péripneumonie contagieuse bovine à des chèvres, Revue d'Elevage et de Medecine Veterinaire des Pays Tropicaux, 53, 319-324.




