- Infectious Diseases of Livestock
- Part 3
- Pasteurellosis in sheep and goats
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Pasteurellosis in sheep and goats
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Pasteurellosis in sheep and goats
W DONACHIE AND M QUIRIE
Introduction
Respiratory disease is common in sheep and goats and is usually the result of a series of complicated interactions involving stress inducers, both physical and physiological, and a variety of infectious agents. The most common form of respiratory disease is pasteurellosis. In lambs and kids affected with the acute disease the signs include fever, dyspnoea, anorexia, recumbence and sudden death. If the animals survive they can become chronically affected with resultant reduced ability to thrive and survive further infections. 2 Pasteurellosis can therefore have a significant impact on the productive efficacy of small stock.
Although the main clinical form of pasteurellosis in sheep and goats is pneumonia there is also a systemic form found only in sheep. The pneumonic form is caused by Mannheimia haemolytica (formerly designated Pasteurella haemolytica1 (see below for more details), and the systemic disease by Bibersteinia trehalosi. In sheep in temperate climates Pasteurella multocida rarely causes pneumonia and little is known of the epidemiology of that infection. Pasteurellosis caused by M. haemolytica is one of the most common bacterial infections of sheep and goats, and by far the most important respiratory one, with a widespread distribution, occurring in temperate, subtropical and tropical climates.7, 11, 15, 21
Aetiology
Pneumonic pasteurellosis in sheep was first described in 19316 but it was not until the 1960s that serotyping and biotyping helped to define the epidemiology of the disease. In sheep, two ‘Pasteurella’ species, M. haemolytica and P. trehalosi, share a common serotyping system comprising a total of 17 serotypes,1, 17, 20 with approximately 90 per cent of all isolates serotypable. Each of the two species, originally biotypes of Mannheimia haemolytica, is associated with a distinct clinical syndrome. M. haemolytica (formerly biotype A) strains are responsible for pneumonic pasteurellosis in sheep and goats of all ages while P. trehalosi strains (formerly biotype T) cause a systemic disease in six to ten-month-old lambs.8
In 1999 the taxonomy of the family Pasteurellaceae changed in response to new information on the relatedness of strains following studies on DNA and ribosomal RNA homology. 1 The revised taxonomy introduced a new genus, Mannheimia, to replace Pasteurella haemolytica and P. haemolytica-like strains. The prototype species of the new genus is Mannheimia haemolytica, which includes all the former P. haemolytica A serotype strains apart from A11. Strains of this latter serotype are now placed in another Mannheimia species,M. glucosida.
The three species are divided into 17 serotypes on the basis of an indirect haemagglutination test, which depends on the serotypes having specific polysaccharide capsules. Serotypes 1, 2, 5, 6, 7, 8, 9, 12, 13, 14, 16 and 17 belong to M. haemolytica, A11 is now M. glucosida and serotypes 3, 4, 10 and 15 areP. trehalosi. The taxonomy of P. trehalosi remains unchanged. The main characteristics of all the new Mannheimia spp. are shown in Table 165.1 and a more detailed list of differences between Mannheimia and Pasteurella spp. is shown in Table 165.2.
A summary of the changes affecting M. haemolytica and P. trehalosi serotyping is shown in Table 165.3.
Mannheimia haemolytica and P. trehalosi are identical in their morphology both being encapsulated, small (1–2mm× 0,3–0,6 mm), Gram-negative, aerobic coccobacilli. In carbohydrate fermentation tests it was originally thought that strains of M. haemolytica fermented arabinose (hence the former biotype A designation) but not trehalose, whereas all P. trehalosi strains ferment trehalose (hence the former biotype T designation). Colonies of M. haemolytica are small and grey with a narrow zone of haemolysis after 24 hours’ incubation. The colonies of P. trehalosi strains are darker, larger (up to 3mm in diameter) and have brownish centres.
The differentiating characteristics of M. haemolytica, P. trehalosi and P. multocida species are shown in Table 165.4.
Table 165.1 Identification of species/taxa within the genus Mannheimia (From Angen et al.1)
| SPECIES | HAEMOLYTICA | GLUCOSIDA | GRANULOMATIS | RUMINALIS | VARIGENA | UNNAMED TAXA | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BIOVARS | A | B | C | D | E | F | G | H | I | 1 | 2 | 1 | 2 | 7A | 8A | 8B | 8C | 9 | 10 | ||
| ODC | − | + | + | + | + | + | − | − | − | + | − | − | − | + | − | − | − | − | − | + | − |
| Arabinose | − | + | − | + | + | − | − | + | − | + | − | − | − | + | + | + | − | − | − | + | + |
| Cellobiose | − | + | + | + | + | + | + | + | + | − | d | − | − | d | − | − | − | − | − | − | − |
| Gentiobiose | − | + | + | − | + | + | + | + | + | − | d | − | − | − | − | − | + | − | − | − | − |
| Inositol | d | + | (+) | + | (+) | (+) | (+) | (+) | (+) | + | d | − | − | d | − | d | (−) | + | + | − | d |
| Maltose | + | + | + | + | + | + | + | + | + | + | d | − | + | d | d | + | + | + | + | + | + |
| Sorbitol | + | + | + | + | + | + | + | + | + | + | + | d | − | − | − | − | − | − | − | + | + |
| Xylose | (+) | + | + | + | + | + | + | + | + | + | d | d | − | + | + | + | + | + | − | + | + |
| NPG | − | + | + | + | + | + | + | + | + | + | + | − | − | d | d | − | − | − | − | − | − |
| ONPF | + | (+) | (+) | + | + | (+) | − | (+) | (+) | + | − | − | − | d | − | − | + | − | − | − | d |
| ONPX | d | + | + | − | − | − | + | − | − | d | (−) | − | − | − | + | d | − | d | − | − | d |
(Characters common to genus: urease −, mannitol +, mannose −, trehalose −)
+ positive, − negative, d positive or negative, (+) positive but some only weakly, (−) negative but some weakly positive
M. haemolytica: Pasteurella haemolytica biogroup 1 (includes all typable strains except A11)
M. glucosida: (biovars A-H), P. haemolytica biogroups 3A-H (includes A11); (biovar I), biogroup 9 (inositol and NPG +ve strains)
M. granulomatis: P. granulomatis, P. haemolytica biogroup 3J and Bisgaard Taxon 20
M. ruminalis: (biovar 1), Bisgaard Taxon 18 (biovars 1 and 3); (biovar 2), P. haemolytica biogroup 8D
M. varigena: (biovar 1), P. haemolytica biogroup 6, Bisgaard Taxon 15 (biovar 1), Bisgaard Taxon 36; (biovar 2), Bisgaard Taxon 15 (biovar 2)
Unnamed taxa: P. haemolytica biogroups 7, 8A, 8B, 8C, 9 (inositol and β-glucosidase -ve strains) and 10
Table 165.2 Characteristics separating Mannheimia and Pasteurella (from Angen et al.1)
| MANNHEIMIA | PASTEURELLA | |
|---|---|---|
| β-haemolysis | − | − |
| α-haemolysis | + | − |
| V-factor dependency | − | d |
| X-factor dependency | − | − |
| VP, 37 °C | − | − |
| Urease | − | d |
| ODC | d | d |
| Indole | −* | +# |
| L-Arabinose | d | − |
| Glucosides | d | − |
| Mannitol | + | d |
| D-Mannose | − | + |
| D-Melibiose | −* | − |
| meso-Inositol | d | − |
| D-Sorbitol | d | d |
| Trehalose | − | d |
| ONPF | d | − |
+ positive, − negative, d positive or negative
* deviating strains occur, # P. avium indole -ve
Bibersteinia trehalosi strains are more resistant than M. haemolytica strains to penicillin, ampicillin, chloramphenicol, oxytetracycline, erythromycin and nitrofurantoin. Basic fuchsin (0,2 mg/ml) in brain-heart infusion broth inhibits M. haemolytica. Methods and reagents used in the characterization of Pasteurella/Mannheimia spp. are:
- Oxidase: use Oxoid touch-sticks (BR64) according to manufacturer’s instructions.
- Catalase: transfer a small amount of growth from blood agar onto a microscope slide using a loop. Add a loopful of catalase reagent (equal volumes of 3 per cent hydrogen peroxide and 1 per cent aqueous methylene blue) and mix. Place a coverslip over the reaction mixture.
- Urease: use Difco urea discs (1625-33-4) according to manufacturer’s instructions (liquid suspension method).
- Indole: inoculate 10 ml nutrient broth, incubate for 48 hours at 37 °C. Take 2ml of broth culture and add 0,1 ml of Kovacs reagent.
- Sugar fermentation tests: Kauffman’s broth base is used. Filter-sterilized sugar solutions (10 per cent) are added to give a final concentration of 1 per cent. Dispense in 3 ml volumes. Prepare a suspension from blood agar in saline to McFarland No. 3–4. Inoculate 200 μl. Incubate for at 37 °C for 14 days. Include sugar-free control. It is recommended that gentiobiose is used as a ‘second stage’ test for only those isolates which require it as it is very expensive. The constituents and preparation of the medium base are:
- Medium base: Bacto beef extract, 5 g; Bacto peptone, 10 g;Na2HPO4, 0,8 g; NaCl, 3 g; bromothymol blue solution (0,04 per cent), 60 ml; and distilled water, 940 ml. Adjust to pH 7,4, autoclave at 121 °C for 15 minutes.
- Rapid detection of glycosidases: Phosphate buffer stock solutions: A, 0.1M monobasic sodium phosphate (15,6 g NaH2PO4.2H2O in 1 000 ml); B: 0.1M dibasic sodium phosphate (14,2 g Na2HPO4 or 35,9 g Na2HPO4.12H2O in 1 000 ml); pH 7,5: 16 ml A + 84 ml B. pH 8,0: 5,3 ml A + 94,7 ml B.
Prepare 0,1 per cent (w/v) solutions of the substrates in the relevant buffer (see Table 165.5) and filter sterilize. Suspend growth from blood agar to at least McFarland No. 5 in 1 ml substrate solution, and incubate at 37 °C for four hours and 24 hours. A positive result is indicated by the development of a yellow colour.
- Ornithine decarboxylation: Prepare decarboxylase broth (Moeller) according to the manufacturer’s instructions with and without L-ornithine. Dispense in 3 ml volumes in bijoux. Prepare a suspension as for the carbohydrate fermentation tests. Inoculate 400:l and overlay with 0,5 to 1 ml of sterile mineral oil. Incubate at 37 °C for seven days.
Table 165.3 Summary of changes in the serotyping of Pasteurella haemolytica
| PREVIOUS DESIGNATION | NEW DESIGNATION |
|---|---|
| P. haemolytica A1, A2, A5-9, A12-14, A16-17 | M. haemolytica (serotypes unchanged) |
| P. haemolytica A11 | M. glucosida A11 |
| P. haemolytica untypable | Most strains M. ruminalis |
| P. trehalosi serotypes T3, T4, T10, T15 | (unchanged) |
Disease associated with Mannheimia haemolytica
The predominant disease caused by this species is pneumonia.
Epidemiology
The prevalence of the serotypes varies (Figure 165.1). In the UK, between 1982 and 1999, 33 per cent of all Mannheimia/ Pasteurella isolates were serotyped as M. haemolytica A2, while A1, A6, A7 and A9 represented about 22 per cent of the total. T10 is the commonest P. trehalosi serotype (11 per cent) followed by T15 (9 per cent) and T4 (7 per cent).
Clinical cases associated with each species have a seasonal distribution with M. haemolytica more prevalent in the spring and summer and P. trehalosi predominant in the autumn. M. haemolytica and P. trehalosi occur in the nasopharynges and tonsils of apparently normal sheep. M. haemolytica predominates in the nasopharynx whereas P. trehalosi is the major species in the tonsils.
Most outbreaks of pneumonic pasteurellosis occur in spring to early summer in temperate regions and many involve both ewes and lambs. Flock outbreaks usually start suddenly with deaths, often in young lambs in which the disease is hyperacute and septicaemic rather than pneumonic.
As lambs get older the disease becomes more confined to the thorax with prominent lesions of pleuritis and pericarditis. In animals beyond three months of age most cases are frankly pneumonic although sudden deaths with septicaemia rather than pneumonia may still occur. As the outbreak progresses over the next few days a number of sheep will be noticed with clinical signs of pneumonia. Observations of the flock show that some sheep have an occasional cough and slight oculo-nasal discharges. Morbidity and mortality rates vary but up to 10 per cent of the flock may be affected.
Pneumonic pasteurellosis also occurs in individual sheep sporadically rather than as part of a clearly defined flock outbreak. In most cases flock outbreaks on individual farms are sporadic and do not occur every year although on some farms small numbers of sheep may succumb annually. There is a tendency for the prevalence of the disease to be higher overall in some years than in others for which there are two possible explanations. Either environmental factors, e.g. climate, are particularly favourable for the disease over a wide area in some years or immunity to the predisposing virus infection (see below) is cyclical.
It is generally assumed that environmental factors are important predisposing causes of pneumonic pasteurellosis but precise epidemiological data are lacking. Some outbreaks can be linked to previous stressful situations such as warm or cold, wet weather, and dipping, castration or dosing.9 There is evidence from experimental investigations and epidemiological studies that infection with parainfluenza type 3 (PI-3) virus, respiratory syncytial virus and sheep pulmonary adenomatosis are factors predisposing to pneumonic pasteurellosis. Infection with PI-3 virus generally produces no or a mild illness and cases of pneumonia in a flock can be attributed to a superimposed infection of the lungs with M. haemolytica strains. Similar to viruses, some respiratory bacterial infections also increase the susceptibility of sheep to secondary M. haemolytica infections. Mycoplasma spp. are common in the respiratory tract of sheep and the combination of Mycoplasma ovipneumoniae and M. haemolytica A2 induces a proliferative subacute or chronic pneumonia in lambs.3 Bordetella parapertussis has recently been isolated from ovine lungs in Scotland and under experimental conditions can predispose to secondary M. haemolytica infections.15
Table 165.4 The differentiation of M. haemolytica, P. trehalosi and P. multocida
| M.(P) HAEMOLYTICA | P. TREHALOSI | P. MULTOCIDA | |
|---|---|---|---|
| Haemolysis | + | + | − |
| Fermentation of xylose | + | − | − |
| Production of indole | − | − | + |
| Growth on MacConkey agar | Small pink colonies | Small pink colonies | − |
Table 165.5 Substrates and buffers for rapid detection of glycosidases
| TEST | ENZYME | SUBSTRATE | pH |
|---|---|---|---|
| NPG | β-glucoside | 4-nitrophenyl-β-D-glucopyranoside (Fluka, 73676) | 8,0 |
| ONPF | α-fucosidase | 2-nitrophenyl-α-L-fucopyranoside (Fluka, 73651) | 7,5 |
| ONPX | β-xylosidase | 2-nitrophenyl-β-D-xylopyranoside (Fluka, 73750) | 8,0 |
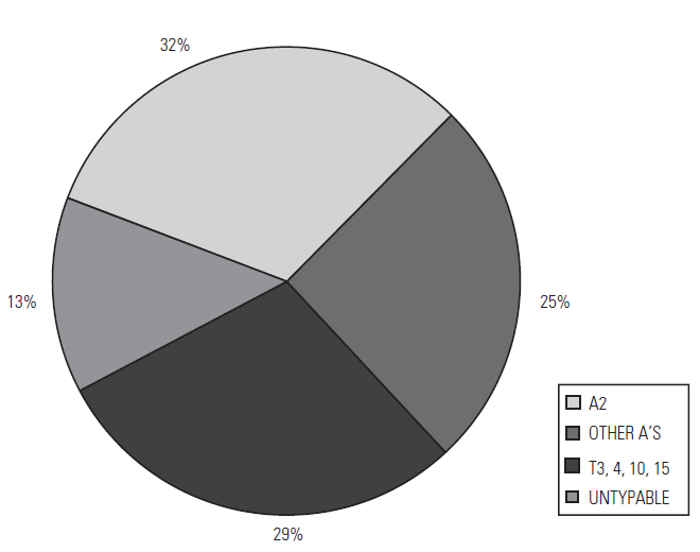
Figure 165.1 Prevalence of M. haemolytica and P. trehalosi serotypes in cases of ovine pasteurellosis in the UK, 1982–1999
Pathogenesis
The lower respiratory tract including the lungs are areas of the body which have to be continually protected against infectious agents. These colonizing agents usually arrive in the nasopharyngeal passages as aerosolized particles in inhaled environmental gases. They then become the source of infectious agents for the lower respiratory tract. The respiratory tract is, however, generally resistant to attacks by airborne pathogens. There exists a dynamic relationship between the innate clearance mechanisms of the host and the impairment of these mechanisms by the pathogens or other factors.
The pathogenesis of pulmonary infections requires a number of stages: colonization of the upper respiratory tract, survival and growth of the agent, and colonization of the lung. In pasteurellosis in sheep and goats the colonization of the upper respiratory tract is regularly achieved as M. haemolytica and P. trehalosi are frequently isolated from apparently healthy animals.9
Mannheimia haemolytica is capable of disrupting the innate defence mechanisms and the integrity of lung tissues in both sheep and goats through a number of mechanisms:
- A specific protease which cleaves IgG1 13
- A specific leukotoxin which attacks the alveolar macrophages, one of the first mechanisms of host defence in the lung19
- Capsular polysaccharide which has been shown to have anti-phagocytic properties.3
Mucociliary clearance is a well-recognized mechanism for removing particles from the lower airways and any conditions that adversely affect its operation will present an opportunity to pathogenic bacteria. Cold temperatures and virus infections are known either to slow down the velocity of the nasal mucus or destroy the epithelial cells in the trachea.2
Viral infection is thought to create an ideal environment consisting of necrotic cells and proteinaceous fluid in the lung favouring bacterial growth by interfering with the mucociliary clearance mechanisms of the respiratory tract, and by depressing the capacity of resident lung macrophages to take up and kill bacteria. The subsequent M. haemolytica infection is exacerbated by these events.
Clinical signs
In a proportion of cases, clinical signs are not noticed and the animal is found dead. In the early stages of flock infection some animals may have a slight, serous, oculonasal discharge with an occasional soft cough. After this more severe clinical signs of the disease occur. The clinical signs of acute, pneumonic pasteurellosis are dullness, anorexia, pyrexia of greater than 40,6 °C and varying degrees of hyperpnoea or dyspnoea. This dyspnoea can be seen as exaggerated movement of the abdominal flank muscles (abdominal lift). On auscultation, adventitious sounds are not prominent and the respiratory sounds are loud and prolonged. There are often serous to mucopurulent nasal and ocular discharges. A frothy fluid drooling from the mouth is usually present in the terminal stages. The morbidity and mortality rates, however, rarely exceed 10 per cent.9 In subacute or chronic cases the clinical signs may be transient and less obvious than in the acute disease. Signs include coughing over a long period, dyspnoea and hyperpnoea which is exacerbated by exercise, dullness and a mucopurulent nasal discharge. 8
Pathology
The initial observation at necropsy of most adult sheep with pneumonic pasteurellosis is extensive ecchymotic haemorrhage in the throat region and over the ribs. On opening the thorax, subpleural and subepicardial petechiation and varying amounts of clear, yellow pleural and pericardial exudate are found.
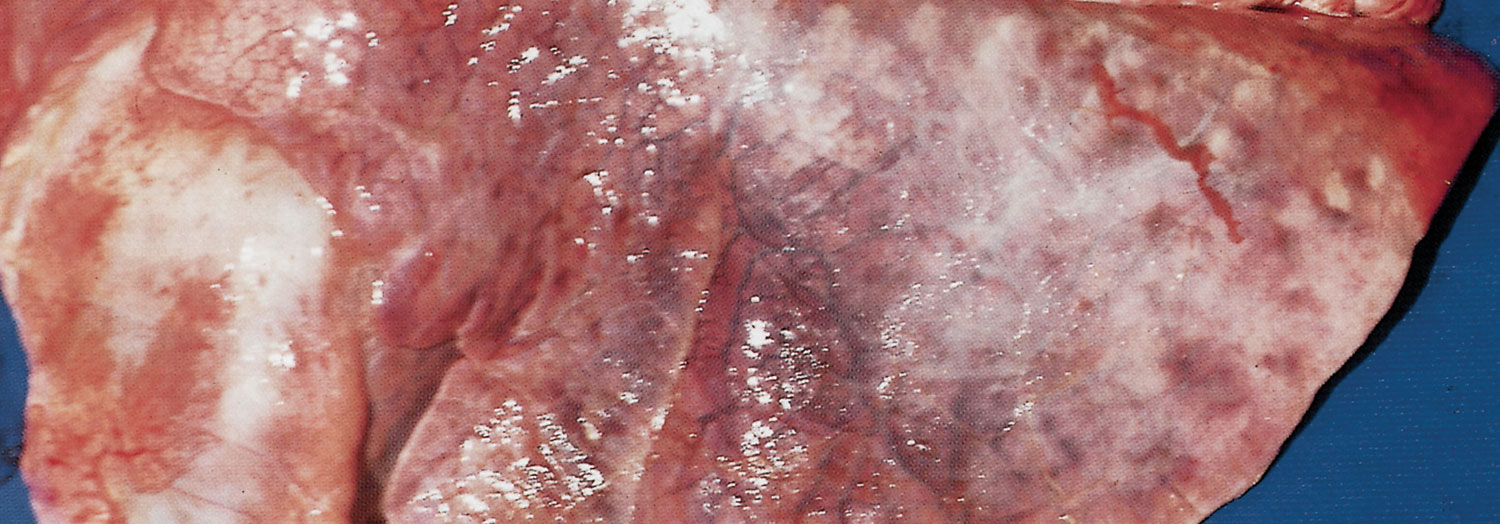
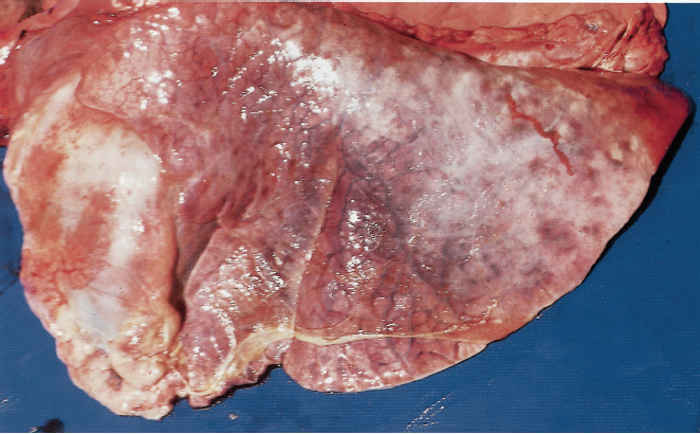
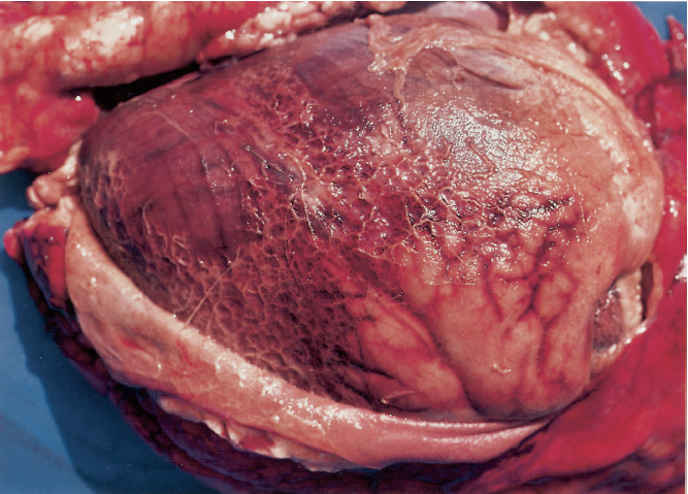
In peracute cases, the lungs are swollen, heavy and cyanotic, with bright purplish-red solid areas which exude a frothy haemorrhagic fluid when incised. In cases of slightly longer duration the lesions are those of an acute fibrinous pleuropneumonia. The lungs are consolidated particularly in their cranial and ventral parts and generally covered by a greenish-yellow, gelatinous pleural exudate (Figure 165.2). The consolidated portions of such lungs contain irregular greenish-brown areas of necrosis, each with a dark, haemorrhagic margin. The tracheobronchial mucosa in peracute and acute pneumonic pasteurellosis is red to dark purple and the airways contain pinkish-stained froth. The pneumonic and pleural lesions are almost invariably accompanied by an acute fribrinous pericarditis (Figure 165.3). In less acute cases greyish-pink, raised areas of consolidation in the cranial lobes predominate which, when incised, resemble liver in appearance and consistency, having red or grey solid lobular tissue prominently separated by thickened septa. In such tissue necrotic foci and an overlaying pleuritis with organizing fibrinous adhesions between the parietal and visceral pleura may be present. Very occasionally the only lesions are solid, raised nodular masses scattered throughout the lungs. These may be mistaken for lung tumours but histology reveals the characteristic pathology of pasteurellosis. The bronchial lymph nodes in pneumonic pasteurellosis are enlarged, soft and pale and may contain numerous petechiae.
In lambs up to 12 weeks of age M. haemolytica is associated with septicaemia rather than a primary pneumonia. There may be either no lung involvement, or pleuritis and pericarditis with only focal areas of pneumonia, pleuritis and pericarditis may be manifested. Petechiae, however, are usually present in the myocardium, spleen, kidneys and liver. The majority of lymph nodes are enlarged and haemorrhagic, and fatty degeneration of the liver is present.
Histologically, in peracute cases the lungs are intensely hyperaemic and haemorrhagic and a pale pink-staining exudate rich in protein mingled with masses of Gram-negative coccobacilli fills the alveoli. The acute consolidated pneumonic lesions are manifested by the presence of a widespread cellular exudate and an accompanying congestion and capillary haemorrhage. Clusters of neutrophils may occur but the predominant cell filling the alveoli is elongated with an intensely basophilic spindle-shaped nucleus called an ‘oat cell’, masses of which form whorls and appear to stream between adjacent alveoli. When present, the characteristic necrotic lesions have a zonal structure with necrosis of all elements centrally surrounded by a dense layer of oat cells, macrophage-like cells and the products of cell death. Progressive stages of pleuritis are observed, the most acute type displays oedema and a diffuse light mononuclear cell infiltration containing masses of coccobacilli, while a thick layer of fibrino-cellular exudate characterizes the less acute form.
Diagnosis and differential diagnosis
Pasteurellosis is the commonest cause of acute pneumonia in sheep, but only a provisional diagnosis should be made on clinical signs and history.8 The presence of M. haemolytica in nasal swabs is of no diagnostic significance and serology is not useful on either an individual or a flock basis. A diagnosis may be provisionally confirmed at necropsy by finding the acute inflammatory changes in the thorax and the characteristic hepatized and/or necrotic lung lesions. Further confirmation may be achieved by histological demonstration of lesions containing oat cells. However, peracute pneumonic pasteurellosis requires bacteriological confirmation, since other conditions, e.g. clostridial disease and even post-mortem autolysis, may cause similarly congested, heavy lungs.
In smears from exudates and from the cut surfaces of lung lesions Gram-negative bacteria may be seen, but only the isolation in culture of large numbers (106 or more colony-forming units [cfu] per g−1 of lung) confirms acute pasteurellosis. In subacute and chronic cases, 103 to 105 cfu g−1 would be expected. Samples for bacteriological examination should be collected from untreated cases and include a range of sites, e.g. thoracic fluids, heart blood, lung lesions, kidney, spleen and liver. Routine serotyping of isolates of M. haemolytica is of little value, but can be useful in epidemiological studies and investigations into putative vaccine breakdowns. Molecular techniques such as pulsed field gel electrophoresis have better discriminatory powers than serotyping and may prove more useful in epidemiological studies.
Similar lesions may be encountered in pneumonia caused by other bacteria, notably P. multocida and rarely Escherichia coli. Aspiration pneumonia may be distinguished to some extent from M. haemolytica pneumonia on the basis of distribution (predilective involvement of the right cranial lobe) and the type of reaction (often gangrenous). Chronic pneumonia due to M. haemolytica should be differentiated clinically and pathologically from jaagsiekte (pulmonary adenomatosis), pneumonia caused by Corynebacterium pseudotuberculosis or Trueperella pyogenes, lungworm infestation and maedi.14 It must, however, be borne in mind that secondary bacterial pneumonia might complicate the clinical and patholgical manifestations of the lung lesions in diseases such as jaagsiekte, maedi and lungworm infestations.
Control
Since pasteurellosis is sporadic, optimum control is achieved by vaccination. However, there are circumstances in which the use of antibiotics is useful, for instance in outbreaks of pasteurellosis in lambs during the period when passive immunity from colostrum has waned and active immunity is being generated by vaccination. Long-acting oxytetracycline has been shown to be effective therapeutically and prophylactically in experimentally infected lambs and under field conditions.
Early commercial vaccines against pasteurellosis were bacterins and contained a very limited range of serotypes. There was no experimental evidence of their efficacy. The development of a method for producing pneumonic pasteurellosis in specific pathogen-free (SPF) lambs allowed the challenge of novel vaccines based on cell extracts and a range of serotypes. It was shown that these extracts were more immunogenic than bacterins and the protection was serotype-specific.
New vaccines based on protein antigens which are involved in iron uptake, have now been developed and became available in 1997. This development was based on the findings that SPF lambs that had recovered from an antibiotic-terminated episode of pasteurellosis induced by exposure to an aerosol of M. haemolytica serotype A2 were solidly immune to challenge with this serotype. This led to the discovery that pasteurellas collected from the pleural fluid (i.e. bacteria grown in vivo) contained proteins not present in pasteurellas grown in vitro and that sheep produced an immune response to those proteins. Identical iron-regulated proteins (IRPs) were produced by the pasteurellas when grown in vitro under conditions of iron limitation and vaccines made from such bacterial cells were highly protective.10 Since IRPs are antigenically similar in all serotypes they offer cross-protection against serotypes not in the vaccine.5 Other antigens are also involved in immunity and leukotoxin-containing vaccines have been shown to be immunogenic. Serological responses in SPF lambs have shown that the leukotoxin-neutralizing titre and the ability of specific antibody and complement to kill M. haemolytica in vitro correlated with immunity to challenge.
Studies on the humoral and cell-mediated immune responses in the sera and lung washings of vaccinated and recovered lambs suggested that humoral immunity was the more important. This was confirmed by the finding that sera or immunoglobulins from infected or hyperimmunized sheep passively protected SPF lambs against aerosol challenge.
The unpredictable, sporadic nature of pasteurellosis necessitates a vaccination pattern that will ensure yearround immunity for all ages of sheep. Breeding sheep should be vaccinated twice at an interval of three to four weeks, around tupping time and given a third, booster dose four to six weeks before lambing to ensure high concentrations of antibodies in the colostrum. Epidemiological evidence suggests that this schedule gives immunity to the lambs up to five weeks of age. Active immunity in lambs should be induced by two doses of vaccine, the first being given as early as ten days after birth as colostrum- acquired antibodies do not interfere with the response to vaccination. Weaned lambs brought in for autumn and winter fattening should be given two doses of vaccine, ideally before arrival on the farm. Annual boosters of breeding stock should be given before lambing but more frequent vaccination can be used to cover periods of high risk as indicated by the pattern of disease on individual farms.
Disease associated with Bibersteinia trehalosi: Systemic pasteurellosis
This is epidemiologically and pathogenetically distinct from the pneumonic form of pasteurellosis.
Epidemiology
Most outbreaks of systemic pasteurellosis conform to a well-defined pattern. In the UK the disease affects six- to nine-month-old sheep during October, November and December. The onset of the disease frequently coincides with folding on rape or turnips or a change to improved pastures and both circumstances have been postulated as predisposing causes. However, there is as yet no proof that they are so. A change to wet, cold weather has also been noted as a possible contributing factor. A typical episode usually starts with a number of sudden deaths, but the number of deaths then quickly drops over the next few days. The mortality rate is quite variable but seldom exceeds 10 per cent of the flock. Sporadic deaths due to systemic pasteurellosis do occur at other times of the year and in all ages of sheep but even less is known about the factors that predispose to these cases.
Successful experimental production of systemic pasteurellosis has been reported. Feedlot lambs given hydrocortisone and a changed diet from 100 per cent roughage (alfalfa) to 90 per cent high protein concentrates succumbed to systemic pasteurellosis.18 Dosing with P. trehalosi was not necessary. It was concluded that the dietary changes induced erosions and ulcers of the gastrointestinal mucosa, which were the portals of entry of the infection leading to the systemic disease. Thus, both epidemiological and experimental evidence points to the same pathogenesis. Pathology similar to that seen in natural causes has also been produced in SPF lambs challenged subcutaneously with P. trehalosi. However, these were young lambs (ten weeks of age) compared with the naturally affected animals (around six to ten months old).12
Clinical signs
The main feature is sudden death, so that affected sheep are seldom seen alive. Those that are, usually are recumbent, extremely depressed, dyspnoeic and frothing at the mouth. This clinical description is consistent with that of endotoxic shock and experimental studies in SPF lambs have demonstrated the typical biochemical characteristics of this condition.12
Pathology
The carcass is usually that of a young sheep in good condition. Subcutaneous haemorrhages are found over the neck and thorax, and ecchymoses are also frequently seen in the pleura and diaphragm, or under the epicardium. The lungs are swollen and oedematous with widespread focal haemorrhages, and frothy blood-tinged fluid exudes from the bronchioles. Consolidation is not a feature. Lesions also occur in the pharynx and upper alimentary tract. In the former site, they take the form of necrotic erosions, which are especially prominent around the tonsillar crypts. Similar erosions may be found also in the nasal mucosa, tongue or soft palate. Necrosis of the oesophageal mucosa, with extensive sloughing of mucosa, may be present and similar necrotic lesions are found variably in the omasum and rumen. The abomasum may contain considerable areas of haemorrhagic inflammation, or shallow haemorrhagic ulcers, most numerous at the pyloric end. Rarely, similar lesions may be seen in the duodenum.
The liver is usually swollen and congested and may contain numerous small (0,5 to 5 mm), grey, necrotic foci scattered throughout its substance. Small infarcts may sometimes be seen in the spleen, and the kidneys appear blotchy. The tonsils and retropharyngeal lymph nodes are usually enlarged and oedematous.
Microscopically, the necrotic lesions in the pharynx and alimentary tract show necrosis of the mucosa with extensive sloughing. Underlying tissues are hyperaemic but exhibit surprisingly little cellular reaction. Masses of Gram-negative coccobacilli and Gram-positive cocci can be seen adhering to the luminal surface of many ulcers or eroded areas and similar masses occlude local vessels and lymphatics. Lesions in the lungs, liver, spleen, adrenals and, less frequently, kidneys can be attributed to the dissemination of bacterial emboli in the terminal arterial system. The lesions are focal and consist of masses of Gram-negative coccobacilli, usually surrounded by zones of necrosis enclosed by basophilic spindle-shaped leukocytes (oat cells). In the brain, serum protein leakage in the cerebrocortical leptomeninges, with mononuclear cell infiltrates in the choroid plexuses of the lateral and fourth ventricles have been reported.8 From the evidence of these findings and the results of experimental work with P. trehalosi in sheep, possible pathogenic mechanisms can be postulated. It is thought that multiplication of P. trehalosi resident in the tonsils occurs under the influence of poorly understood environmental factors, e.g. change in pasture, with the development of necrotizing lesions in the pharynx and upper alimentary tract. Bacterial emboli from these sites pass by way of the general circulation to the lungs and other organs, where further multiplication and toxin release cause the death of the animal. An alternative hypothesis is that the primary lesions are in the forestomachs and intestines with emboli passing to the lungs via the liver and portal system.
Diagnosis
The diagnosis of systemic pasteurellosis depends on the isolation in culture of large numbers (>106 cfu g−1 of tissue) of P. trehalosi from the lungs, livers and spleens of sheep with the gross pathological changes described. Bibersteinia trehalosi can be isolated from the nasopharynges and tonsils of apparently normal sheep and in small numbers from other sites including the lungs but their presence in these sites can be ignored. It should also be remembered that antibiotic therapy may reduce the numbers of bacteria in the lesions.
Control
Because of the epidemiology of the disease, control is best achieved by vaccination. As with pneumonic pasteurellosis, field trials are difficult due to the sporadic nature of the disease on individual farms from year to year. Experimental studies in SPF lambs have demonstrated that vaccines containing iron-regulated proteins such as those used in M. haemolytica conferred significant protection against T10 and T15 challenges.5
Most isolates of P. trehalosi are sensitive to oxytetracycline but as sheep are rarely seen in the early stages of the disease, therapy is seldom possible and there are no reports on the value of prophylactic therapy.
Since stress may play some part in predisposing to this disease, flock management should be designed to minimize the stress involved in changes of environment and nutrition during the late autumn and early winter.
Other Mannheimia haemolytica and Bibersteinia trehalosi infections
Mannheimia haemolytica is the cause of other pathological conditions but these occur less frequently than pneumonic and systemic pasteurellosis. Strains of M. haemolytica and P. trehalosi have been isolated from cases of mastitis in ewes and this condition is occasionally fatal. Arthritis is a common sequel to experimental intravenous inoculation of P. trehalosi and is also occasionally diagnosed in the field. Sporadic Mannheimia/Pasteurella meningitis affects ewes and lambs. Diagnosis of all these conditions depends on the isolation of large numbers of Mannheimia/Pasteurella cells from the lesions.
References
- ANGEN, O., MUTTERS, R., CAUGANT, D.A., OLSEN, J.E. & BISGAARD. M., 1999. Taxonomic relationships of the [Pasteurella] haemolytica complex as evaluated by DNA-DNA hybridisations and 16S rRNA sequencing with proposal of Mannheimia haemolytica gen. nov., comb. nov., Mannheimia granulomatis comb. nov., Mannheimia glucosida sp. nov., Mannheimia ruminalis sp. nov. and Mannheimia varigena sp. nov. International Journal of Systematic Bacteriology, 49, 67–86.
- BROGDEN, K.A., LEHMKUHL, H.D. & CUTLIP. R.C., 1998. Pasteurella haemolytica complicated respiratory infections in sheep and goats. Veterinary Research, 29, 233–254.
- CZUPRYNSKI, C.J., NOEL, E.J. & ADLAM, C., 1991 Interaction of bovine alveolar macrophages with A1 in vitro: Modulation by purified capsular polysaccharide. Veterinary Microbiology, 26, 349–358.
- DONACHIE, W. & JONES, G.E., 1982. The use of ELISA to detect antibodies to Pasteurella haemolytica A2 and Mycoplasma ovipneumoniae in sheep with experimental chronic pneumonia. In: wardley, r.c. & crowther, j.r., (eds). The ELISA: Enzyme-linked immunosorbent assay in veterinary research and diagnosis. Current Topics in Veterinary Medicine and Animal Science. Vol. 22. The Hague, Boston, London: Martinus Nijhoff.
- DONACHIE, W., 1995. Vaccine development against Pasteurella haemolytica infections in sheep. In: donachie, w., lainson, a. & hodgson, c., (eds). Haemophilus, Actinobacillus and Pasteurella. New York: Plenum Publishing Company.
- DUNGAL, N., 1931. Contagious pneumonia in sheep. Journal of Comparative Pathology, 44, 126–143.
- FODOR, L., VARGA, J. & HATJOS, I., 1989. Characterization of Pasteurella haemolytica isolated from goats in Hungary. Acta Veterinaria Hungarica, 37, 35–38.
- GILMOUR, N.J.L. & GILMOUR, J.S., 1985. Diagnosis of pasteurellosis in sheep. In Practice, 7, 145–149.
- gilmour, n.j.l. & gilmour, j.s., 1989. Pasteurellosis of sheep. In: adlam, c. & rutter, j.m., (eds). Pasteurella and Pasteurellosis. London: Academic Press.
- GILMOUR, N.J.L., DONACHIE, W., SUTHERLAND, A.D., GILMOUR, J.S., JONES, G.E. & QUIRIE, M., 1991. Vaccine containing iron-regulated proteins of Pasteurella haemolytica A2 enhances protection against experimental pasteurellosis in lambs. Vaccine, 9, 137–140.
- HAYASHIDANI, H., HONDA, E., NAKAMURA, T., MORI, Y., SAWADA, T. & OGAWA, M., 1988. Outbreak of pneumonia caused by Pasteurella haemolyticainfection in Shiba goats in Japan. Japanese Journal of Veterinary Science, 50, 960–962.
- HODGSON, J.C., MOON, G.M. QUIRIE, M. & DONACHIE, W. 1993. Biochemical signs of endotoxaemia in lambs challenged with T10 strain of Pasteurella haemolytica and the effect of vaccination on the host response. Proceedings of the Sheep Veterinary Society, 17, 201–204.
- LEE, C.W & SHEWEN, P.E., 1996. Evidence of bovine immunoglobulin IgG1 (IgG1) protease activity in partially purified culture supernate of Pasteurella haemolytica A1. Canadian Journal of Veterinary Research, 60, 127–132.
- NESBIT, J.W., BASTIANELLO, S.S. & BATH, G., 1993. Ovine and caprine pasteurellosis. In: coetzer, j.a.w., thomson, g.r. & tustin, r.c., (eds). Infectious Diseases of Livestock with Special Reference to Southern Africa. Cape Town: Oxford University Press Southern Africa.
- ODENDAAL, M.W. & HENTON, M.M., 1995. The distribution of Pasteurella haemolytica serotypes among cattle, sheep and goats in South Africa and their association with disease. Onderstepoort Journal of Veterinary Research, 62, 223–226.
- PORTER, J.F., CONNOR, K. KREUGER, N., HODGSON, J.C. & DONACHIE, W, 1995. Predisposition by an ovine isolate of Bordetella parapertussis to subsequent infection with Pasteurella haemolytica A2 in specific pathogen free lambs. Journal of Comparative Pathology, 112, 381–389.
- SNEATH, P.H.A. & STEVENS, M., 1990. Actinobacillus rossii sp. nov., Actinobacillus seminis sp. nov., nom. rev., Pasteurella bettii sp. nov., Pasteurella lymphangitidis sp. nov., Pasteurella mairi sp. nov., and Bibersteinia trehalosi sp. nov. International Journal of Systematic Bacteriology, 40, 148–153.
- SUAREZ-GUEMES, F., COLLINS, M.T. & WHITEMAN, C.E., 1985. Experimental reproduction of septicaemic pasteurellosis in feedlot lambs: Bacteriologic and pathological examinations. American Journal of Veterinary Research, 46, 185–192.
- SUTHERLAND, A.D., 1985. Effects of Pasteurella haemolytica cytotoxin on ovine peripheral blood leucocytes and lymphocytes obtained from gastric lymph. Veterinary Microbiology, 10, 431–438.
- YOUNAN, M. & FODOR, L., 1995. Characterisation of a new P. haemolytica serotype (A17). Research in Veterinary Science, 58, 98.
- ZAMRI-SAAD, M., SUBRAMANIAM, P., SHEIK-OMAR, A.R., SANI, R.A. & RASEDEE, A., 1994. The role of concurrent haemonchosis in the development of pneumonic pasteurellosis in goats. Veterinary Research Communications, 18, 119–112.




