- Infectious Diseases of Livestock
- Part 3
- Clostridium perfringens type D infections
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Clostridium perfringens type D infections
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Clostridium perfringens type D infections
N P J KRIEK, M W ODENDAAL AND P HUNTER
Introduction
Pulpy kidney disease is a peracute, acute or occasionally chronic infectious enterotoxaemic disease that is common in sheep, uncommon in goats, and virtually unknown in cattle in South Africa. It is caused by the systemic effects of Clostridium perfringens type D epsilon toxin which is absorbed into the general circulation from the intestinal tract where it is elaborated by the rapidly multiplying organisms. Pulpy kidney disease in sheep affects animals of any age, but young animals in good condition are particularly susceptible. Predisposing factors, such as sudden changes in diet from poor to good quality, precipitate the occurrence of the disease. Peracute and acute cases are characterized by sudden death, and chronic cases by the development of a neurological syndrome that results from focal symmetrical encephalomalacia (FSE). In goats, the disease may be peracute, as in sheep, or it may manifest as an acute or chronic diarrhoea due to enterocolitis2 terminating in death, or as a chronic, lingering and debilitating condition.4 The differences between the disease in sheep and in goats, in general, relate to clinical signs and responses to treatment and vaccination. 4
Pulpy kidney disease occurs all over the world wherever small ruminants are maintained. It was first described by Gilruth in New Zealand in 1903 and thereafter in Australia, the UK, the USA and South Africa. The association of an unknown potent toxin with the pathogenesis of this disease was first demonstrated by Montgomerie and Rowlands and then by other workers in 1931, 1933 and 1934. The organism, then referred to as Bacillus ovitoxicus, was isolated in 1932.25 The bacterium, now known as C. perfringens type D, occurs in large numbers in the intestinal tract of normal sheep without causing any clinical signs and can be regarded as an obligate parasite.6, 55
In South Africa it is believed that many of the cases diagnosed as ‘geilsiekte’ (prussic acid poisoning) in the past were, in fact, pulpy kidney disease.53 That the disease remains one of the most important diseases of sheep in South Africa today is confirmed by the annual vaccine sales which exceed 50 million doses.
Aetiology
Clostridium perfringens type D has the same morphological and biochemical characteristics as the other members of the perfringens group and produces two major lethal toxins, alpha and epsilon, and a number of minor toxins, which may be either lethal or non-lethal (Clostridium perfringens group).
Epsilon toxin, the significant major toxin and cause of pulpy kidney disease, is produced as a prototoxin.32, 39, 40, 51, 69, 71, 72 Its in vitro production is dependent on the presence of suitable conditions which must include optimal pH and the availability of an appropriate carbohydrate source.23, 24
Epidemiology
Pulpy kidney disease is one of the most important diseases of sheep in southern Africa, as it is in many other parts of the world. It occurs in sheep of all ages, but most commonly in lambs of two age groups; from three weeks to three months, and from 6 to 12 months of age. Cases of C. perfringens type D enterotoxaemia have been reported in seven- to ten-week- old calves,21 in young cattle in good condition up to two years old,30 as well as in adult animals.37 Though cattle of all ages are affected, it is mainly a disease of calves and young cattle up to two years of age in feedlots. In the USA, C. perfringens type D is associated with overeating disease or sudden death of feedlot cattle. As opposed to the disease in sheep, it only affects individual animals over a period.30 In goats, most outbreaks have been reported in dairy animals3although it has also been described in Angora goats under extensive conditions in South Africa.73 The condition in goats appears to be poorly understood and has not been properly defined. Significant differences occur between pulpy kidney disease in goats and sheep.63
In endemic areas, C. perfringens type D is harboured in the alimentary tract of animals as an obligate parasite and its presence in this situation provides the focus for a potentially fatal infection. Small numbers occur in the forestomachs, but not in the abomasum. The numbers increase towards the ileum where most C. perfringens organisms are found. Conditions here are ideal for the organisms to multiply under normal conditions. The numbers become less in the large intestine, but sufficient numbers of the organism are excreted in the faeces to allow its bacteriological isolation. As is the case with type B, type D isolates are not considered part of the microbial soil flora, as they cannot compete with type A isolates in this habitat. Clostridium perfringens type D is, however, able to survive for a number of months in soil before dying.55, 57
There appears to be a number of predisposing factors which precipitate the occurrence of the disease. These include stasis of the intestinal tract; sudden changes in the feeding regimen; sudden changes from poor to lush pastures; grazing on fodder crops, such as green oats, green wheat and green lucerne (alfalfa); very high protein diets; overeating on concentrates and lush pastures;26 deworming; and coccidiosis.41 Sudden changes in the weather and wilting of pasture grasses have also been recorded as playing predisposing roles. Under experimental conditions pulpy kidney disease is induced within 25 hours of infusion of C. perfringens type D cultures.4 For sheep this period may vary from two to five days under natural conditions.
Focal symmetrical encephalomalacia may occur either as a sporadic disease involving one or a small number of sheep, or in outbreak form. Sheep of all ages are affected, although it is often the case that when FSE does make its appearance in a flock, two specific age groups of animals are particularly affected: weaner lambs up to the age of 10 to 12 weeks, and mature sheep, three years of age or older.19 The factors which predispose to FSE are the same as those in pulpy kidney disease, cases often being preceded by a change to a lush pasture or by anthelmintic drenching. Focal symmetrical encephalomalacia is more likely to occur in flocks with a partial immunity to C. perfringens type D. The interval between exposure to the predisposing factor and the appearance of clinical signs is usually 7 to 14 days, but it may be as short as two days.
Pathogenesis
Clostridium perfringens type D is present as a commensal in the lumen of the intestinal tract and proliferates profusely when conditions in the intestinal lumen become suitable for it to do so, such as those which may arise when the diet of a ruminant is suddenly changed from a poor to rich one, or if an animal engorges on palatable feed. Ruminal flora become adapted to the diet that the animal is eating, and if the diet is changed, a period lasting several days is required for the flora to readapt to the new conditions. The ruminal flora of an animal consuming veld grass consist of a high proportion of cellulolytic organisms. If its feed is suddenly changed to one with a high starch content, such as one of the cereals, particles of starch, which would be converted to low chain fatty acids in the rumen of an animal adapted to its diet, pass unchanged from the rumen into the intestine. The starch is used by the saccharolytic C. perfringens type D, which multiplies rapidly and elaborates large amounts of epsilon toxin,6 as well as its other toxins.
The appearance of starch granules in the small intestine mayincrease the viable colony count of C. perfringens type D to as many as 109 organisms per gram of intestinal content (with a correspondingly high concentration of epsilon toxin) within a few hours.7 Epsilon toxin is generally produced in small amounts in the intestines of carrier animals of C. perfringens type D, but these amounts are too small to engender any deleterious effects although they can stimulate antibody formation. It has been shown that up to 54 per cent of animals in an unvaccinated flock may contain serum epsilon antitoxin titres.3
Epsilon toxin is produced in the small intestines as a virtually non-toxic prototoxin which is activated by trypsin and chymotrypsin, but not by pepsin, in the intestinal tract. Activation increases the toxicity by at least a thousand fold. 6, 45 Small amounts of the epsilon toxin in the intestinal tract have no effect on the intestinal mucosa, but large concentrations increase its permeability so that within a number of hours large amounts of toxin are absorbed into the systemic circulation.6 The mu toxin of C. perfringens type D also increases the permeability of the mucosa. Epsilon toxin is endotheliotoxic and, after attaching to binding sites on capillary endothelial cells to form large complexes, remains associated with the cell membrane to induce the development of large pores.50 The consequence of this cell injury is a rapid loss of intracellular K+ and increased levels of intracellular Na+ and Cl−. Further progression of cell injury is associated with increased intracellular concentrations of Ca++50 and cell death.54 Systemically absorbed epsilon toxin causes an increased permeability of capillaries in many organs, including those in the intestinal mucosa, thus further enhancing the absorption of the toxin.46
Epsilon toxin selectively binds to the luminal surface of the endothelium of capillaries of the brain and, to a lesser extent, of other organs, such as those in the kidneys that surround the loops of Henle and the distal convoluted tubules and the hepatic sinusoids.11 The extracellular oedema that develops in the brain is the result of the breakdown of the tight junctions of the vascular endothelium, and is responsible for the main clinical signs of pulpy kidney disease,11 particularly the neurological signs that are manifested by the acute cases. It appears that mechanisms other than those associated with oedema are involved in the development of nervous signs seen in pulpy kidney disease. The neurotoxicity induced in mice injected intravenously with epsilon toxin was associated with excessive glutamate release in the hippocampus that led to the development both of neuronal damage and of seizures; cerebral ischaemia that may be the consequence of endothelial injury and extensive oedema appeared not to play a role in the development of these lesions. 43 In the brains of rats intoxicated subacutely with epsilon toxin, neuronal damage thought to be the consequence of a direct cytotoxic effect, cause the development of progressive neuronal cytoplasmic vacuolation and shrunken, hyperchromatic cells with marked nuclear pyknosis.15
In FSE, the lesions in the brain progress sequentially, within a period of seven days, from an initial vasogenic oedema to the development of bilaterally symmetrical malacic foci that principally involve the white matter of certain areas of the brain.14
The majority of animals suffering from pulpy kidney disease become hyperglycaemic and glycosuric, but the precise pathogenetic mechanisms responsible for this development remain unresolved. Several hypotheses have, however, been suggested, such as glycogenolysis of hepatic reserves, stimulation of cAMP by hepatocyte-bound epsilon toxin, the release of catecholomines (which activate cAMP) due to the brain oedema,8, 9, 18, 70 or by reflex hypersecretion of adrenalin from the adrenal medulla in response to oedema of the hypothalamus.
The reason why goats respond to the effects of the toxin in a different way from sheep is not known.4 The lesions in the brain of goats suffering from natural and experimental pulpy kidney disease reflect the presence of, respectively, severe microangiopathy and an associated perivascular proteinaceous exudation, and severe vasogenic oedema.62, 65 Although lesions have been detected in the brain of goats suffering both from experimental and natural pulpy kidney disease, larger doses of alpha toxin appear to be required to induce these changes.65
Clinical signs
The course of the acute form of the disease in lambs is very short—often less than two hours and very rarely more than 12 hours.5 Many animals are, however, found dead without any premonitory signs of illness being noticed. Usually the first indication of the disease in a flock is the finding in the morning of sheep that have died overnight.
When affected animals are closely observed, the first transient signs to be noticed include dullness, depression, facial movements and loss of appetite. Signs preceding death indicate severe neurological derangement and include hypersensitivity, listlessness, hyperaesthesia, staggering gait, knuckling of the fetlocks, ataxia progressing to recumbency, clonic convulsions, opisthotonus, marked salivation, stertorous respiration between convulsions, and terminal coma.11, 26 The body temperature is usually normal but may become elevated if the convulsions are severe. Young lambs, three to four weeks old, may suffer from colic and manifest abdominal bloating before death supervenes. 36 Manifestations of the disease in adult sheep are similar to those in lambs, but the course of the disease in some may be as long as 24 hours.
The clinical signs of FSE vary according to the acuteness of the disease and involve only those which indicate impairment of the central nervous and alimentary systems. Two different syndromes are seen: an acute neurological disorder, the so-called stagger syndrome in which sheep are unable to stand and with death occurring within 24 hours; and a more chronic condition, the ‘dummy syndrome’, in which the sheep can stand but show general weakness or paresis when handled.19 In the field, animals suffering from the latter syndrome usually die within seven days of developing clinical signs, but they may linger for up to 14 days. If nursed they may survive for a longer period. Animals suffering from either of these syndromes have no consistent abnormalities in body temperature or respiratory or heart rates.
An animal suffering from the acute syndrome of FSE is usually found in lateral recumbency and comatose but, when stimulated, shows normal or exaggerated motor responses. It appears to be blind, lacks the eye preservation reflex, and when handled, develops opisthotonus. Spontaneous convulsions are rare. Nystagmus is seen in about a quarter of affected animals. The rumen is usually atonic and the faeces invariably soft and pasty.19
Animals suffering from the ‘dummy syndrome’ lose their fear of dogs and humans, wander around aimlessly, and manifest head-pressing and teeth-grinding. Severely affected sheep lack fine muscular co-ordination and are blind. The sheep are inappetent and show paralysis of the muscles of prehension, thus losing their ability to eat and drink. There is usually ruminal stasis and the faeces are pasty and unformed. When lying, they assume a position of sternal recumbency, which is also the position in which they die.19
Three syndromes of C. perfringens type D enterotoxaemia have been reported in goats. The peracute or ‘sudden death’ syndrome is similar to that in sheep. The acute syndrome is characterized by an acute diarrhoea and signs of abdominal discomfort, often culminating in recovery or death two to four days after the onset of clinical signs. The chronic syndrome may persist for days or weeks and is characterized by diarrhoea and progressive weight loss.3, 4
Clostridium perfringens type D enterotoxaemia in cattle generally occurs in the form of sporadic outbreaks. The peracute disease is characterized by sudden death, and the acute by nervous signs and a subnormal body temperature. The animals appear dull and listless, and wander around aimlessly. There are signs of colic and salivation, and when they become prostrate, they assume a position of lateral recumbency and manifest opisthotonus. They show depression, anorexia and appear to be blind before expiring or recovering completely.21, 37, 44
Pathology
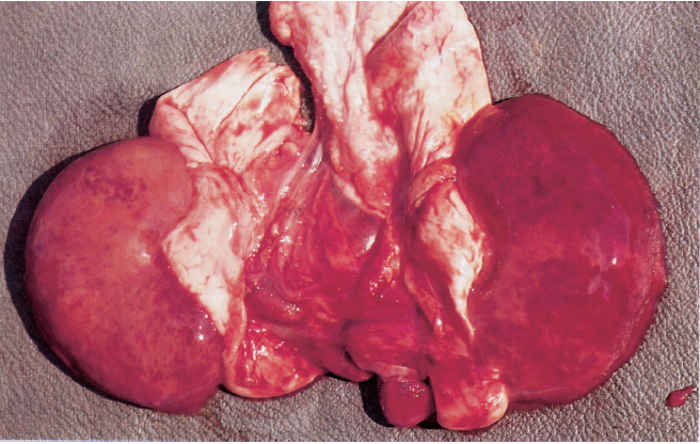
Sheep that have died from pulpy kidney disease are usually in good condition. The rapid onset of post-mortem changes is characteristic, particularly those in the kidneys, which develop more quickly than in other organs, the rapid autolysis soon reducing the renal cortices to a pulpy mass17, 26 (Figure 180.1).
Lesions in sheep dying from the peracute to acute disease are not constant and may be virtually absent in some. When present, one or more of the following may be observed to some degree: varying numbers of petechial or ecchymotic haemorrhages in the subcutis of the dorsal region of the neck, shoulders and rump; petechial or ecchymotic haemorrhages in the parietal peritoneum, muscles of the flank, the muscular portion of the diaphragm, epicardium and endocardium, especially that of the left ventricle, and thymus; an excessive amount of fluid in the pericardial sac, which clots partially when exposed to air; congestion and oedema of the lungs; distension of the small intestine, which may be patchily congested and contain gas (even in a ‘fresh’ carcass); mild to moderate nephrosis and partial (‘blotchy’) to complete congestion of the renal cortices, (the degree of post-mortem changes present in the kidneys may be gauged by comparing them to those in the liver; in addition, these changes may result in the post-mortem degeneration and rupture of the walls of small blood vessels and the consequent presence of small, scattered, cortical haemorrhages — the renal congestion and haemorrhages giving rise to the Afrikaans name of ‘bloednier’ (literally, ‘blood kidney’) for the disease); and glycosuria which may disappear with the passage of time due to the post-mortem growth of bacteria in the urine.
In the majority of cases there is no macroscopic evidence of enteritis in animals dying peracutely, although intestinal congestion is present in some cases. In very young lambs dying of pulpy kidney disease, the abomasum may rupture terminally.36
The most prominent haematological and biochemical changes occurring in lambs after the parenteral administration of epsilon toxin include: haemoconcentration (anhydraemia), metabolic acidosis, hyperglycaemia, glycosuria, and an elevation of the blood concentrations of pyruvate, lactate and alpha-ketoglutarate as a result of the increase in circulating glucose.16
In goats that have died of the peracute disease, lesions may be similar to those in sheep. In the acute to chronic disease a severe pulmonary oedema and changes in the intestinal tract are the most consistent lesions. A hydropericardium containing a fibrin clot similar to that in sheep is not always present in these animals.4 The lesions in the intestinal tract are those of a fibrinous to fibrinonecrotic enterocolitis. The intestinal content may be green or redtinged and mucoid and may in some instances contain pure blood.22 The presence of mucofibrinous casts in the lumen of the large intestine is a constant finding2 and numerous ulcers may occur in the intestinal mucosa.63 Mesenteric lymph nodes are swollen and oedematous.
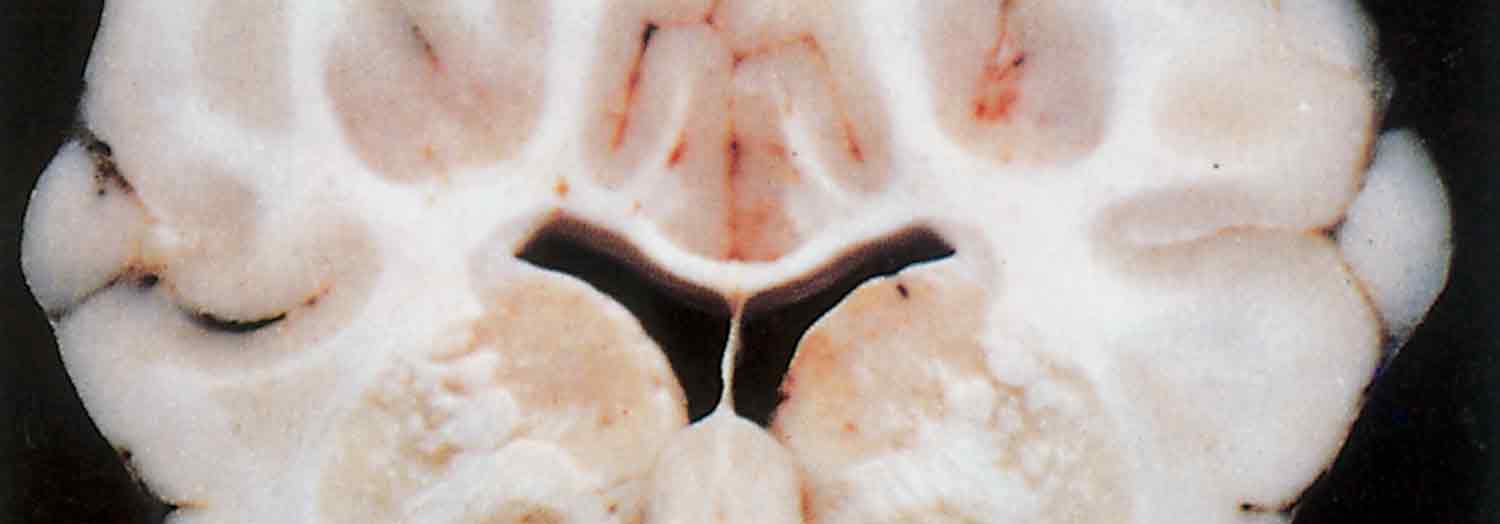
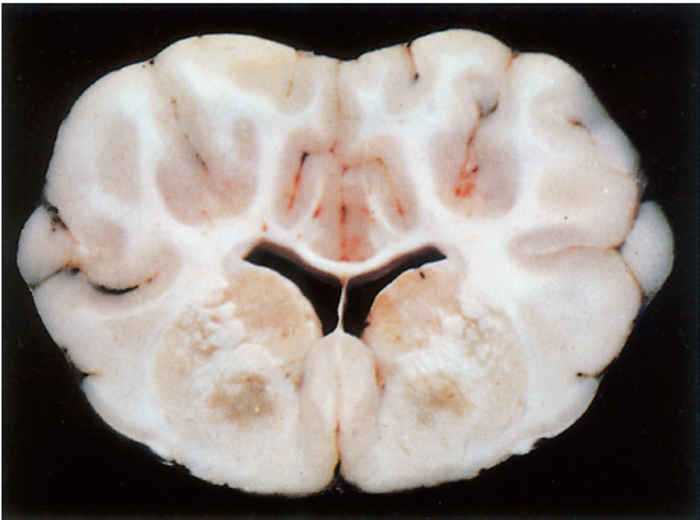
In sheep that have died from FSE, the only lesions consistently present, although they are frequently microscopic, occur in the brain. Those that have suffered from the acute FSE syndrome may also manifest some of the lesions present in pulpy kidney disease. Lesions in the brain are bilaterally symmetrical and two general patterns of distribution are recognized; areas of leukoencephalomalacia may be present in the internal capsule, dorsolateral thalamus, substantia nigra, basal ganglia and cerebellar peduncles,19 the most constant sites being the thalamus and the midbrain areas11 or the white matter of the frontal lobes. Areas of malacia are better seen in formalin-fixed brain tissue.
Three basic histologic lesions occur in the brain of sheep affected by FSE.10 These are widespread perivascular oedema, perivascular haemorrhage and malacia (Figure 180.2). Areas of malacia are always focal and sharply demarcated, and are often accompanied by haemorrhage, but lack an inflammatory reaction.10 Ultrastructurally, there is swelling of astrocytes, the perivascular oedema being caused by their swollen end-feet, and severe damage to vascular endothelium is apparent.11
No consistent clinical pathological changes are encountered in sheep suffering from FSE. In some, however, hyperglycaemia and glycosuria may be present.19
Lesions in cattle closely resemble those of pulpy kidney disease in sheep.
Differential diagnosis
Diseases and conditions known to cause sudden death, and in particular other clostridial enterotoxaemias, such as that caused by Clostridium sordellii,52 should be considered as differential diagnoses. Diseases of particular importance in the differential diagnosis of pulpy kidney disease include heartwater; bluetongue; Rift Valley fever; the plant poisonings leading to sudden death, such as ‘gousiekte’ (‘quick death’) due to Pachystigma pygmaeum and certain other members of the family, ‘gifblaar’ (‘poison leaf’) poisoning caused by ingestion of Dichapetalum cymosum, which contains monofluoroacetate as the toxic principal, and plants containing cyanogenetic glycosides which are responsible for prussic acid poisoning (‘geilsiekte’ in Afrikaans); nutritional myopathy (white muscle disease); and ruminal acidosis due to the overeating of grains.
Listeriosis and nutritional myopathy in calves, milk fever in cows and acute lead poisoning may be confused with enterotoxaemia. 44 Focal symmetrical encephalomalacia may be confused clinically with cases of hypocalcaemia, copper deficiency, colisepticaemia, pregnancy toxaemia, coenurosis, cerebrocortical necrosis, cranial abscessation, botulism and tick paralysis.
Diagnosis
A presumptive diagnosis of C. perfringens type D enterotoxaemia may be based on the clinical signs, lesions and history as well as on the demonstration of the presence of epsilon toxin and C. perfringens typeDin the intestinal content. The use of a specific polymerase chain reaction (PCR) to detect the presence of genes encoding the toxins (alpha, epsilon and beta) of Clostridium perfringens in formalin-fixed, paraffin- embedded intestinal tissues of sheep and goats can now also be used to confirm the presence of the bacteria. This method is very specific and highly sensitive and correlates well with the results of conventionally culturing the bacteria.68 Confirmation of the diagnosis is, however, not always easy as the causative organisms are normal commensals in the intestinal tract and epsilon toxin may occur in the intestinal lumen of animals immune to the toxin but which have died from other diseases. In addition, failure to demonstrate the presence of the toxin in a typical case does not necessarily preclude the diagnosis of pulpy kidney disease. 10
Additional evidence for the confirmation of the disease may be obtained by histological examination of the brain of sheep. Even though signs of malacia may only be macroscopically present in about 30 per cent of cases, histologically the combined presence of severe oedema, haemorrhages and malacia in the brain stem and midbrain areas is highly suggestive of pulpy kidney disease and is considered by some to be the most reliable way of diagnosing enterotoxaemia in sheep.19 The malacic areas are invariably focal and bilaterally symmetrical.10
Demonstration of low serum antibody levels on a flock basis may provide further supporting evidence for the diagnosis. Epsilon antitoxin levels exceeding 0,15 units/ml virtually exclude a diagnosis of pulpy kidney disease.
Appropriate specimens should be submitted for confirmation of the diagnosis of pulpy kidney disease (see the introduction, Clostridium perfringens group).
In order to determine the immune status of the flock, submit individual samples each consisting of 10 ml of serum from at least 10 different animals.
Specimens from dead animals should be taken as soon as possible after their death. As putrefactive changes interfere with diagnostic procedures and may preclude the making of a diagnosis, those for bacteriological and toxicological examination should be collected by using aseptic techniques, and they, as well as the samples of urine, effusions and serum samples, should immediately be refrigerated or packed in ice and kept in the cold state until received by the diagnostic laboratory.
Several in vitro immunological assays for the detection of epsilon toxin in specimens are available. A reverse phase, passive haemaglutination assay (RPHA) detects concentrations of epsilon toxin as low as 6 × 10−7 mg/ml, whereas the single radial immunodiffusion technique is only capable of detecting epsilon toxin above the level of 0,01 mg/ml.1 Counter-immunoelectrophoresis is more effective than the older mouse protection assay for the detection of epsilon toxin in the intestinal contents.29 A latex agglutination test has been developed for the detection of epsilon toxin of C. perfringens type D in the intestinal contents of animals suspected of dying from enterotoxaemia.42 An ELISA assay has also been developed as an alternative to the mouse neutralization test. It is faster, giving qualitative results in four hours, and is more sensitive than the mouse test,45 but cross-reactions with other clostridial toxins may complicate the interpretation of the results of this assay. The ELISA techniques used to determine the antibody levels against C. perfringens beta and epsilon tocoid antigens have become more refined with the use of monoclonal antibodies and show a significant correlation with the mouse neutralization test and are used in potency assays of these antigens in mono- or multivalent vaccines.13
For the diagnosis of FSE, the whole brain of the affected animal should be preserved in an adequate quantity of 10 per cent formalin solution for histopathological examination.
Control
Animals affected by pulpy kidney disease invariably die too acutely to allow any form of treatment.
The simultaneous administration of a hyperimmune serum to epsilon toxin and a Clostridium perfringens type D oil adjuvant vaccine to three-month-old susceptible lambs, has been found to give passive protection for 14 days without any interference occurring in the development of an active immunity.47
Managemental practices should be altered to reduce those conditions which are known to predispose to C. perfringens type D enterotoxaemia. In lambs, tail-docking, castration and marking should be performed at an early age so as not to cause a reduction in feed and milk intake, as these are conducive to the development of the condition.36 It is also to be recommended that managemental practices be so designed that sudden changes in the diet of animals, particularly from a poor to a carbohydrate-rich one, or overeating, are prevented. If such changes can be anticipated, the animals should be vaccinated prior to the event.3 It may be advisable to vaccinate before drenching with anthelmintics if a flock’s immune status is in doubt.
Natural exposure of sheep and goats to epsilon toxin does not, with few exceptions, stimulate the induction of serum antitoxin titres of sufficient magnitude that will provide protection against the effects of the toxin when it becomes elaborated in the intestinal tract in such quantities that overt disease is precipitated.3, 20 Effective control of the disease cannot be accomplished without the application of immunoprophylactic measures. Adequate serum levels of epsilon antitoxin give complete protection against C. perfringenstype D enterotoxaemia. Serum antibody levels of at least 0,15 international units of epsilon antitoxin/millilitre are sufficient to protect sheep against the experimentally induced disease.31
An alum-precipitated C. perfringens type D (pulpy kidney) epsilon toxoid vaccine is used to protect lambs from pulpy kidney disease.27, 33, 59 Good immunity is induced by two vaccinations at a month’s interval but it is short-lived, lasting only 15 weeks.34 Emulsifying the epsilon toxoid in complete or incomplete Freund’s adjuvant induces the development of an adequate level of immunity which persists for two years after a single injection,34 but this formulation unfortunately gives rise to unacceptable tissue reactions at the site of inoculation. This problem was overcome by formulating an oil emulsion vaccine in which Arlacel A and Tween 80 were used as emulsifiers.60 This formulation was most effective when tested in a multicomponent vaccine, containing all the clostridial antigens of veterinary importance to be found in adult sheep.61 Vitamin E ([dl]-atocopherol acetate) has also been used successfully as an adjuvant when mixed with C. perfringens typeDepsilon toxoid in weaned lambs.58 A number of multicomponent clostridial vaccines are available which, inter alia, protect against pulpy kidney disease.
Young lambs between 2 and 44 days of age do not respond well to immunization against pulpy kidney disease.28, 56 The presence of colostrum-derived passive immunity in lambs born of immune dams protects them for 13 to 19 weeks35 and does not interfere with their ability to respond to active immunization.47, 48 The vaccine regimen in lambs two to three months of age should consist of the administration either of two doses of an oil adjuvant vaccine at an interval of four weeks, or of an oil adjuvant vaccine followed four weeks later by an alum-precipitated vaccine. This gives rise to a good immune response which remains effective for at least six to eight months. An alum-precipitated vaccine given as a booster at this stage results in an immunity which lasts for at least a year.12 Annual immunization thereafter with an alum-precipitated vaccine should provide sufficient protection against C. perfringens type D enterotoxaemia. Animals vaccinated for the first time against the disease show serum antibody titres after a period of 10 to 14 days.38 Adult animals vaccinated for the first time are given the oil adjuvant vaccine followed four to six weeks later by the alum- precipitated vaccine. This is repeated after six to eight months, thereafter annually.
In order to provide young lambs with a colostrum-derived passive immunity which will protect them until they are old enough to be actively immunized (i.e. older than 45 days), pregnant ewes may be vaccinated. Ewes which have been previously inoculated require only a single vaccination four weeks before lambing, while those which have not been previously immunized should be vaccinated twice, at an interval of four to six weeks, the second injection being given about four weeks before lambing.
The antibody response of goats to vaccination is similar to that of sheep, but in spite of the development, following repeated vaccinations, of serum antibody levels which should be protective, the disease itself is not effectively controlled. The administration of the vaccine to goats may, in addition, give rise to the development of persistent lumps at the site of the injection, which may be confused with caseous lymphadenitis lesions.3 The efficacy of the C. perfringens epsilon toxoid in incomplete Freund’s adjuvant and in aluminium hydroxide adjuvant under experimental conditions has been demonstrated.66
A new development in biotechnology has led to the production and use of an anti-idiotypic antibody against the epsilon toxin of Clostridium perfringens type D. This anti-idiotypic antibody recognizes variable-region determinants on specific antigen-binding antibodies. It has been demonstrated that this anti-idiotypic antibody can be used to generate an immune response against the epsilon toxin in animals which have not been exposed to the toxin.49
References
- BEH, K.J. & BUTTERY, S.H., 1978. Reverse phase passive haemagglutination and single radial immunodiffusion to detect epsilon antigen of Clostridium perfringens type D. Australian Veterinary Journal, 54, 541–544.
- BLACKWELL, T.E. & BUTLER, D.G., 1992. Clinical signs, treatment, and post mortem lesions in dairy goats with enterotoxaemia: 13 cases (1979– 1982). Journal of the American Veterinary Medical Association, 200, 214–217.
- BLACKWELL, T.E., BUTLER, D.G. & BELL, J.A., 1983. Enterotoxaemia in the goat: The humoral response and local tissue reaction following vaccination with two different bacterin-toxoids. Canadian Journal of Comparative Medicine, 47, 127–132.
- BLACKWELL, T.E., BUTLER, D.G., PRESCOTT, J.F. & WILCOCK, B.P., 1991. Differences in signs and lesions in sheep and goats with enterotoxaemia induced by intraduodenal infusion of Clostridium perfringens type D. American Journal of Veterinary Research, 52, 1147–1152.
- BLOOD, D.C. & HENDERSON, J.A., 1968. Veterinary Medicine. 3rd edn. London: Bailliére, Tindall & Cassell.
- BULLEN, J.J., 1970. Role of toxins in host-parasite relationships. In: kadis, s., montie, t.c. & ajl, s.j., (eds). Microbial Toxins. Vol. IIA, 233–275. New York & London: Academic Press.
- BULLEN, J.J. & SCARISBRICK, R., 1957. Enterotoxaemia of sheep: Experimental reproduction of the disease. Journal of Pathology and Bacteriology, 73, 495–509.
- BUXTON, D., 1978. Further studies on the mode of action of Clostridium welchii type D epsilon toxin. Journal of Medical Microbiology, 11, 293–298.
- BUXTON, D., 1978. In vitro effects of Clostridium welchii type D epsilon toxin on guinea pig, mouse, rabbit and sheep cells. Journal of Medical Microbiology, 11, 299–302.
- BUXTON, D., LINKLATER, K.A. & DYSON, D.A., 1978. Pulpy kidney disease and its diagnosis by histological examination. The Veterinary Record, 102, 241.
- BUXTON, D. & MORGAN, K.T., 1976. Studies of lesions produced in the brains of colostrum deprived lambs by Clostridium welchii (C. perfringens) type D toxin. Journal of Comparative Pathology, 86, 435–447.
- CAMERON, C.M., 1980. Effective immunization of lambs against enterotoxaemia. Onderstepoort Journal of Veterinary Research, 47, 287–289.
- EBERT, E., ÖPPLING, V., WERNER, E. & CUSSLER, K., 1999. Development and prevalidation of two different ELISA systems for the potency testing of Clostridium perfringens and toxoid containing veterinary vaccines. FEMS Immunology and Medical Microbiology, 24, 299–311.
- FINNIE, J.W., 1984. Histopathological changes in the brain of mice given Clostridium perfringens type D epsilon toxin. Journal of Comparative Pathology, 94, 363–370.
- FINNIE, J.W., BLUMBERG, P.C. & MANVIS, J., 1999. Neuronal damage produced in rat brains by Clostridium perfringens type D epsilon toxin. Journal of Comparative Pathology, 120, 415–420.
- GARDNER, D.E., 1973. Pathology of Clostridium welchii type D enterotoxaemia. I. Biochemical and haematological alterations in lambs. Journal of Comparative Pathology, 83, 499–507.
- GARDNER, D.E., 1973. Pathology of Clostridium welchii type D enterotoxaemia. II. Structural and ultrastructural alterations in the tissues of lambs and mice. Journal of Comparative Pathology, 83, 509–524.
- GARDNER, D.E., 1973. Pathology of Clostridium welchii type D enterotoxaemia. III. Basis of the hyperglycaemic response. Journal of Comparative Pathology, 83, 525–529.
- GAY, C.C., BLOOD, D.C. & WILKINSON, J.S., 1975. Clinical observations of sheep with focal symmetrical encephalomalacia. Australian Veterinary Journal, 51, 266–269.
- GRINER, L.A., 1961. Enterotoxaemia of sheep. III. Clostridium perfringens type D antitoxin titers of normal, nonvaccinated lambs. American Journal of Veterinary Research, 22, 447–448.
- GRINER, L.A., AICHELMAN, W.W. & BROWN, G.D., 1956. Clostridium perfringens type D (epsilon) enterotoxaemia in Brown Swiss dairy calves. Journal of the American Veterinary Medical Association, 129, 375–376.
- HARWOOD, D.G., 1996. Clostridial disease: A diagnostic dilemma. Goat Veterinary Society Journal, 16, 26–30.
- HAUSCHILD, A.H.W., 1966. Selective effect of pH on the production of exocellular protein by Clostridium perfringens type D. Journal of Bacteriology, 92, 800–801.
- HAUSCHILD, A.H.W. & PIVNICK, H., 1965. Effect of carbohydrates on toxigenesis by Clostridium perfringens type D. Canadian Journal of Microbiology, 11, 15–22.
- HENNING, M.W., 1949. Animal Diseases in South Africa. 2nd edn. Pretoria: Central News Agency.
- HENNING, M.W., 1956. Animal Diseases in South Africa. 3rd edn. Pretoria: Central News Agency.
- HEPPLE, J.R., CHODNICK, K.S. & PRICE, E.K., 1959. Immunization of lambs against Clostridium welchii type D enterotoxaemia (pulpy kidney disease) with a purified toxoid, aluminium treated. The Veterinary Record, 71, 201–207.
- HOEFLER, W.C. & HALLFORD, D.M., 1985. Growth response and survival rates of lambs immunized against enterotoxaemia and/or treated with enterotoxaemia antotoxin. Agri-Practice-Ovine Medicine, 6, 22–25.
- HORNITSKY, M., ROMALIS, L.F., ROSS, A.D. & SHELDRAKE, R.F., 1989. Comparison of counterimmunoelectrophoresis with mouse protection assay in the detection of epsilon toxin in the intestinal contents of goats. Australian Veterinary Journal, 66, 121–122.
- HUNGERFORD, T.G., 1990. Diseases of Livestock. 9th edn. Sydney: McGraw-Hill Book Company.
- JANSEN, B.C., 1960. The experimental reproduction of pulpy kidney disease. Journal of the South African Veterinary Medical Association, 31, 205–208.
- JANSEN, B.C., 1965. Enterotoxaemia: An immunological study. DSc thesis. Potchefstroom University for a Christian Higher Education.
- JANSEN, B.C., 1967. The production of a basic immunity against pulpy kidney disease. Onderstepoort Journal of Veterinary Research, 34, 65–80.
- JANSEN, B.C., 1967. The duration of immunity to pulpy kidney disease of sheep. Onderstepoort Journal of Veterinary Research, 34, 333–344.
- JANSEN, B.C., 1976. The standardization of Cl. perfringens antigens and antisera. Developments in Biological Standardization, 32, 35–44.
- JOUBERT, J.P., VERMEULEN, S.O. & KELLERMAN, G.E., 1987. Bloednier in die suiplam (in Afrikaans). Journal of the South African Veterinary Association, 58, 43.
- KEAST, J.C. & MCBARRON, E.J., 1954. A case of bovine enterotoxaemia. The Australian Veterinary Journal, 30, 305–306.
- KENNEDY, K.K., NORRIS, S.J., BECKENHAUER, W.H. & WHITE, R.G., 1977. Vaccination of cattle and sheep with a combined Clostridium perfrigens types C and D toxoid. American Journal of Veterinary Research, 38, 1515–1517.
- KULSHRESTHA, S.B., 1974. Effect of carbohydrates on the production of lethal toxin by Clostridium perfringens types B and C with particular reference to beta toxin and epsilon toxin. Indian Journal of Animal Science, 43, 628–631.
- KULSHRESTHA, S.B., 1974. Effect of period of incubation and pH on the production of beta and epsilon toxins by Clostridium welchii type B and type C. Indian Journal of Animal Science, 43, 987–990.
- MARSH, H. & TUNNICLIFF, E.A., 1944. Enterotoxaemia in feedlot lambs in connection with an outbreak of coccidiosis. Journal of the American Veterinary Medical Association, 104, 13–14.
- MARTIN, P.K. & NAYLOR, R.D., 1994. A latex agglutination test for the qualitative detection of Clostridium perfringens epsilon toxin. Research in Veterinary Science, 56, 259–261.
- MIYAMOTO, O., SUMITANI, K., NAKAMURA, T., YAMAGAMI, S., MIYATA, S., ITANO, T., NEGI, T. & OKABA, A., 2000. Clostridium perfringens epsilon toxin causes excessive release of glutamate in the mouse hippocampus. FEMS Microbiology Letters, 189, 109–113.
- MUMFORD, D.H., 1961. Enterotoxaemia in cattle. The Australian Veterinary Journal, 37, 122–126.
- NAYLOR, R.D., MARTIN, P.K. & SHARPE, R.T., 1987. Detection of Clostridium perfringens epsilon toxin by ELISA. Research in Veterinary Science, 42, 255–256.
- NIILO, L., 1986. Enterotoxemic Clostridium perfringens. In: gyles, c.l. & thoen, c.o., (eds). Pathogenesis of Bacterial Infections in Animals. Ames, Iowa: Iowa State University Press.
- ODENDAAL, M.W., VISSER, J.J., BERG, N. & BOTHA, W.J.S., 1989. The effect of passive immunization on active immunity against Clostridium perfringens type D in lambs. Onderstepoort Journal of Veterinary Research, 56, 251–255.
- OXER, D.T., MINTY, D.W. & LIEFMAN, C.E., 1971. Vaccination trials in sheep with clostridial vaccines with special reference to passively acquired Clostridium welchii type D antitoxin in lambs. Australian Veterinary Journal, 47, 134–140
- PERCIVAL, D.A., SHUTTLEWORTH, A.D., WILLIAMSON, E.D. & KELLY, D.C., 1990. Anti-idiotypic antibody-induced protection against Clostridium perfringens type D. Infection and Immunity, 58, 2487–2492.
- PETIT, L., MAIER, E., GIBERT, M., POPOFF, M.R. & BENZ, R., 2001. Clostridium perfringens epsilon toxin induces a rapid change of cell membrane permeability to ions and forms channels in artificial lipid bilayers. Journal of Biological Chemistry, 276, 15736–15740.
- PIVNICK, H., HAUSCHILD, A.H., GORENSTEIN, B. & HABEEB, A.F, 1965. Effect of controlled pH on toxigenesis by Clostridium perfringens type D. Canadian Journal of Microbiology, 11, 45–55.
- POPOFF, M.R., 1984. Bacteriological examination in enterotoxaemia of sheep and lamb. The Veterinary Record, 114, 324.
- SCHULZ, K.C.A. & MCINTYRE, C., 1948. Preliminary report on an infectious enterotoxaemia-like disease in sheep. Journal of the South African Veterinary Medical Association, 19, 93–99.
- SHORT, S.J., TITBAL, R.W. & LINDSAY, C.D., 2000. An assessment of the in vitro toxicology of Clostridium perfringens type D epsilon-toxin in human and animal cells. Human and Experimental Toxicology, 19, 108–116.
- SMITH, L.D.S., 1975. The Pathogenic Anaerobic Bacteria. 2nd edn. Springfield, Illinois: Charles C. Thomas.
- SMITH, L.D.S. & MARSH, H., 1953. The immunization of young lambs against enterotoxaemia. American Journal of Veterinary Research, 14, 408–410.
- SMITH, L.D.S. & WILLIAMS, B.L., 1984. The Pathogenic Anaerobic Bacteria. 3rd edn. Springfield, Illinois: Charles C. Thomas.
- TENGERDY, R.P., MEYER, D.L., LAUERMAN, L.H., LUEKER, D.C. & NOCKELS, C.F., 1983. Vitamin E enhanced humoral antibody response to Clostridium perfringens type D in sheep. British Veterinary Journal, 139, 147–151.
- THOMSON, A. & BATTY, I., 1953. The antigenic efficiency of pulpy kidney disease vaccines. The Veterinary Record, 65, 659–663.
- THOMSON, R.O. & BATTY, I., 1967. Experimental clostridial oil emulsion vaccines. Bulletin de L’Office International des Epizooties, 67, 1569–1581.
- THOMSON, R.O., BATTY, I., THOMSON, A., KERRY, J.B., EPPS, H.B.G. & FOSTER, W.H., 1969. The immunogenicity of a multicomponent clostridial oil emulsion vaccine in sheep. The Veterinary Record, 85, 81–85.
- UZAL, F.A., GLASTONBURY, J.R., KELLY, W.R. & THOMAS, R., 1997. Caprine enterotoxaemia associated with cerebral microangiopathy. The Veterinary Record, 141, 224–226.
- UZAL, F.A., & KELLY, W.R., 1996. Enterotoxaemia in goats. Veterinary Research Communications, 20, 481–492.
- UZAL, F.A. & KELLY, W.R., 1997. Effects of the intravenous administration of Clostridium perfringens type D epsilon toxin on young goats and lambs. Journal of Comparative Pathology, 116, 63–71.
- UZAL, F.A. & KELLY, W.R., 1998. Protection of goats against experimental enterotoxaemia by vaccination with Clostridium perfringens type D epsilon toxoid. The Veterinary Record, 142, 722–725.
- UZAL, F.A. & KELLY, W.R., 1998. Experimental Clostridium perfringens type D enterotoxaemia in goats. Veterinary Pathology, 35, 132–140.
- UZAL, F.A., PASINI, M.I., OLAECHEA, F.V., ROBLES, C.A. & ELIZONDO, A., 1994. An outbreak of enterotoxaemia Clostridium perfringens type D in goats in Patagonia. The Veterinary Record, 135, 279–280.
- WARREN, A.L., UZAL, F.A., BLACKALL, L.L. & KELLY, W.R., 1999. PCR Detection of Clostridium perfringens type D in formalin-fixed, paraffin wax-embedded tissue of goats and sheep. Applied Microbiology, 29, 15–19.
- WORTHINGTON, R.W. & MÜLDERS, M.S.G., 1979. Isolation and characterization of antibodies to Clostridium perfringens epsilon toxin from hyperimmune horse serum. Onderstepoort Journal of Veterinary Research, 46, 121–124.
- WORTHINGTON, R.W., BERTSCHINGER, H.J. & MÜLDERS, M.S.G., 1979. Catecholamine and cyclic nucleotide response of sheep to the injection of Clostridium welchii type D epsilon toxin. Journal of Medical Microbiology, 12, 497–502.
- WORTHINGTON, R.W., MÜLDERS, M.S.G. & VAN RENSBURG, J.J., 1973. Clostridium perfringens type D epsilon prototoxin. Some chemical, immunological and biological properties of a highly purified prototoxin. Onderstepoort Journal Veterinary Research, 40, 145–152.
- WORTHINGTON, R.W., MÜLDERS, M.S.G. & VAN RENSBURG, J.J., 1973. Enzymatic activation of Clostridium perfringens epsilon prototoxin and some biological properties of activated toxin. Onderstepoort Journal Veterinary Research, 40, 153–156.
- VAN TONDER, E.M., 1975. Notes on some disease problems in Angora goats in South Africa. Veterinary Medical Reviews, 1/2, 109–138.




