- Infectious Diseases of Livestock
- Part 3
- Eperythrozoonosis
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Eperythrozoonosis
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
Eperythrozoonosis
F T POTGIETER
Introduction
Eperythrozoonosis is caused by Eperythrozoon spp., which are rickettsial organisms that occur epicellularly on erythrocytes and thrombocytes, as well as freely in the plasma of a wide variety of animal species including pigs, sheep, cattle, dogs, cats, mice and rats. Most infections are subclinical, but varying degrees of fever, anaemia and icterus occur (particularly in immunocompromised or splenectomized animals). Natural cases of eperythrozoonosis are generally not of great clinical or economic significance in livestock.
The genus Eperythrozoon was established in 1928 by Schilling75 when he named the parasites he observed in mouse blood Eperythrozoon coccoides.5 In 1934 Neitz et al.63 were the first to describe Eperythrozoon ovis in sheep in South Africa. In this article a note by Neitz and Quinlan was added, which reported that they had also seen an Eperythrozoon sp. in splenectomized calves. In the same year, Adler and Ellenbogen1 described Eperythrozoon wenyoni, which they had observed in blood smears of a splenectomized calf.
Hoyte36 found and named Eperythrozoon teganodes, an organism that occurs free in the blood of cattle. He also suggested that some highly pleomorphic species of Eperythrozoon may eventually prove to be mixed infections of more than one species. Although many scientists have recognized E. teganodes,7, 24, 26, 27, 37, 50, 99 this organism is not listed in Bergey’s Manual of Systematic Bacteriology.48
A third species in cattle, which seems to exclusively infect thrombocytes, was first reported as occurring in Madagascar9 and later in Finland.7, 98 Uilenberg100 named this organism Eperythrozoon tuomii, and it was subsequently concluded that this parasite is a valid species.105 All three species in cattle have been identified in Argentina and in Germany.22, 26 However, E. tuomii, which also occurs in South Africa, does not seem to have earned taxonomic status, even after it was shown to have no antigenic relationship to E. wenyoni.97
Eperythrozoon wenyoni, E. teganodes and E. tuomii are generally regarded as being non-pathogenic for cattle. There is, however, a report of fatal eperythrozoonosis occurring in five- and six-month-old calves.6
Eperythrozoonosis of pigs and sheep are probably the only two diseases of veterinary importance in livestock to be caused by eperythrozoa.24 According to Jansen,43 an eperythrozoon had been identified in blood smears from pigs in Zaire by P.J. du Toit of the Onderstepoort Veterinary Institute in South Africa as early as 1942. In the USA it was found that a condition of pigs associated with icterus and anaemia was caused by an eperythrozoon.86 Splitter 84 named this parasite Eperythrozoon suis and also described another non-pathogenic variety, Eperythrozoon parvum. The latter parasite was also described in pigs in South Africa by Jansen.44 Studies in splenectomized pigs revealed that E. parvum appears to be more pathogenic than had been originally thought.3 Although E. suis is regarded as an economically significant parasite in pigs, it is considered to be unimportant in the relatively small pig industry in southern Africa.
The aetiology of eperythrozoonosis in sheep in South Africa is well documented.59, 63 Extensive reviews have been published on eperythrozoonosis and haemobartonellosis comparing the aetiology and other aspects of these infections.24, 46, 47
Eperythrozoon ovis may play a role in the poorly understood ill-thrift syndrome (Ill thrift) in sheep and goats particularly in the Eastern Cape Province of South Africa.
Eperythrozoon spp. do have a nuisance value in splenectomized donors of live-blood vaccines and in experimental animals. Subclinical Eperythrozoon spp. infections in such animals may result in the occurrence of aberrations in experimental data,21 probably due to the reactivation of latent infections. For example, sheep dosed with Pachystigma spp. plants, the cause of ‘gousiekte’, often develop clinical eperythrozoonosis.21
* For recent changes in the nomenclature of these organisms, refer to the introduction to Section 3, Rickettsial and chlamydial diseases
Aetiology and life cycle
Eperythrozoa are non-motile, prokaryotic, parasitic microorganisms belonging to the order Rickettsiales, family Anaplasmataceae. Morphological differentiation between some Eperythrozoon and Haemobartonella spp. is difficult.24, 47 Depending on the species, these parasites occur epicellularly on erythrocytes and thrombocytes, as well as freely in the blood plasma. Their association with the erythrocyte membrane is used to differentiate between some species.
A summary of the morphological characteristics and location in the blood of the different organisms occurring in domestic stock in South Africa is given in Table 44.1. The characteristic ring forms of Eperythrozoon spp. seen in thin blood smear preparations stained with Giemsa are probably artefacts formed by the spherical organisms during the drying process.24, 45, 46 The proportions of the morphological types of E. ovis apparently change as the parasitaemia increases. 25 Both scanning and transmission electron microscopy have added considerably to our knowledge of their morphology, multiplication and association between them and the cells they parasitize.25, 45, 52, 81, 98, 102, 105
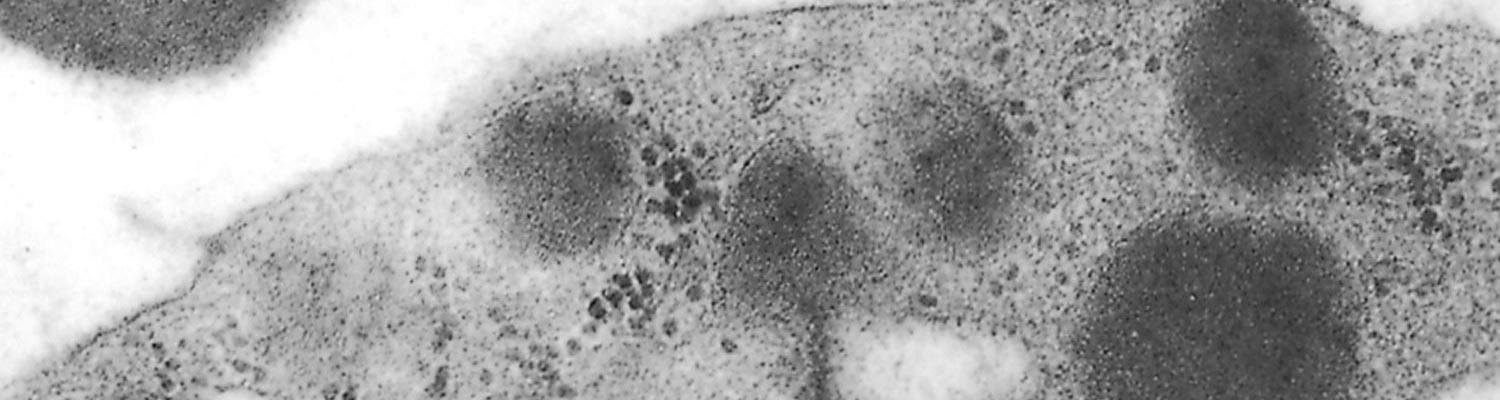
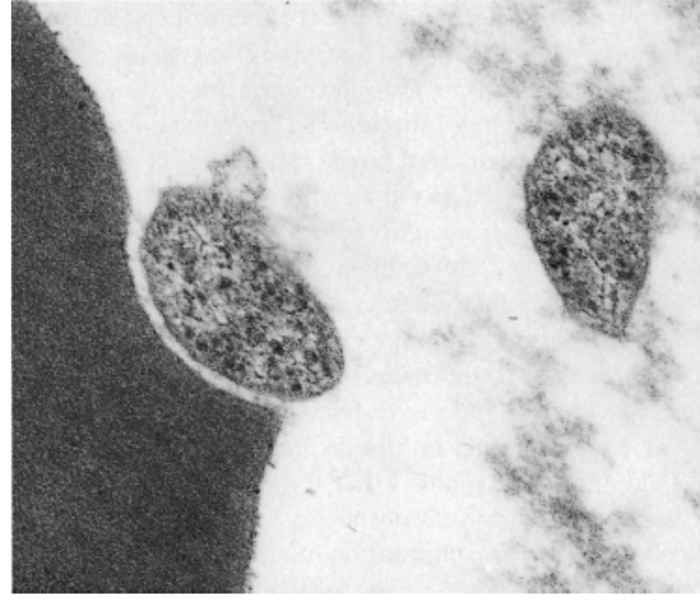
There is little difference between the fine structure of eperythrozoa25, 52, 69, 98, 102, 105 and Haemobartonella bovis.82 The organisms consist of a single membrane enveloping an amorphous matrix containing small ribosome-like granules. Vacuoles and delicate filaments are also sometimes seen within the matrix. Most of these organisms seem to be separated from the host cell membrane by a definite space (Figure 44.1 and Figure 44.2).
In the case of E. ovis, rod- and spherical-shaped parasites are situated in depressions of the surface of erythrocytes. There are varying degrees of intimacy in the association between the parasite and erythrocyte.25 Eperythrozoon suis, although closely associated with the erythrocyte, is separated from the plasmalemma by a definite space,102 the interaction between parasitesandthe cells results in severe deformation of the erythrocyte membrane. Eperythrozoon ovis is attached to the red blood cell by a thread-like structure,39 whereas E. wenyoni, which seems to lie in a slight depression on the surface of the erythrocyte, has a loose relationship with the cell and the parasite apparently causes little harm to its host.45
It appears that E. suis replicates in that small immature forms bud from larger immature and mature forms which seem to correspond to the coccoid, discoid and ring forms of E. suis.102 Eperythrozoon ovis also multiplies by budding, and in this way, chains or clumps of organisms are formed in animals with a severe parasitaemia.25 Scanning electron microscope studies on E. wenyoni showed that the parasites occur alone or in chains, clusters, or pairs, which suggests that both budding and binary fission may take place.45
It has been suggested that the bone marrow may be a primary site for the multiplication of E. ovis and that morphologic differences are due to the stage of multiplication.39
The complete life cycle of Eperythrozoon spp. is still unknown.
Epidemiology
Eperythrozoon spp. are very successful parasites, being widely distributed throughout the world, causing little disease and surviving indefinitely in the host.
Table 44.1 Eperythrozoon spp. that infect domestic stock in southern Africa
| PARASITE | HOST | LOCALITY | SIZE/DIAMETER | MORPHOLOGY IN GIEMSA-STAINED SMEARS* |
|---|---|---|---|---|
| E. wenyoni | Cattle | Epi-erythrocytic and free | 0,3–1,5 μm | Rings (typical), discs and coccoids are common forms which stain reddish-purple. Parasites form clusters around circumference of erythrocytes |
| E. teganodes | Cattle | Free in plasma | Rods and threads 0,3–3,5 μm (length) 0,2–0,3 μm (width) | Slender rods and threads are most common. Rings with rod or thread attached (frying-pan outline) are rare, but characteristic of species. Stain reddish-purple |
| E. tuomii | Cattle | Attached to thrombocytes, occasionally free in plasma attached to granulocytes (exceptional) | Rings 0,4–0,5 μm | Ring, ovoid and coccoid forms. Darkly stained rods are occasionally seen. Parasites completely surround thrombocytes in severe infections |
| E. ovis | Sheep, goats | Epi-erythrocytic and free in plasma | 0,5–1,0 μm | Rings, rods and irregularly shaped parasites may cover or form clusters around circumference of red blood cells. Stain pale purple or pinkish purple |
| E. suis | Pigs | Epi-erythrocytic and occasionally free in plasma | Normal 0,8–1,0 μm Large forms 0,3 μm | Ring forms predominate (large ring forms have distorted appearance). Coccoid, disc and rod forms also occur. Stain pale-purple to reddish-purple |
| E. parvum | Pigs | Epi-erythrocytic and free in plasma | 0,5–0,8 μm | Coccoid and occasional ring forms. Parasites accumulate in large numbers on individual red blood cells. Stain pale-purple to reddish-purple |
* The colour varies with the pH of the buffer and the batch of stain
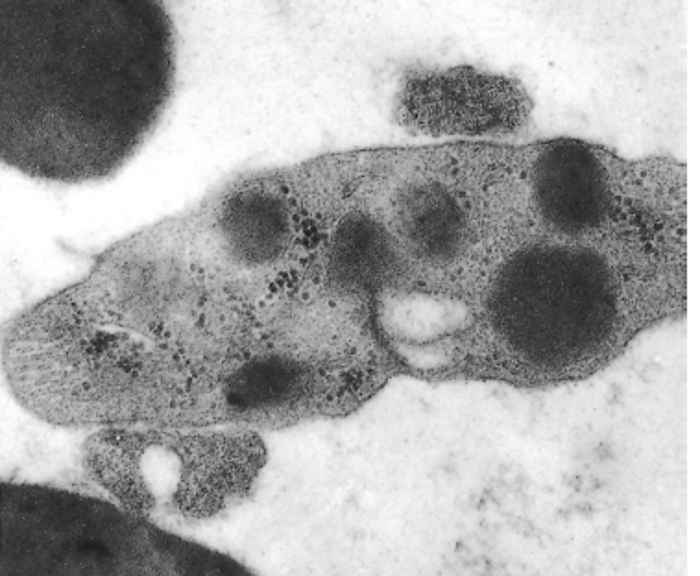
A variety of Eperythrozoon spp. infect sheep, goats, cattle and pigs in southern Africa (Table 44.1) and are probably very prevalent. However, very little information is available on their epidemiology in the subcontinent. It is believed that heartwater, babesiosis and anaplasmosis live-blood vaccines, produced inadvertently from Eperythrozoon carriers, unintentionally caused the widespread distribution of this organism in southern Africa.70
In Germany it has been shown in a survey of blood smears from 570 pigs that 5,7 per cent of healthy pigs harboured E. suis.31
Little is known about the natural transmission of Eperythrozoon spp. It is generally believed that they are arthropod-borne; the pig louse (Haematopinus suis) is capable of transmitting E. parvum43 and certain mosquitoes (Culex annulirostris and Aedes camptorhynchus) can transmit E. ovis mechanically.13, 34 Stable flies (Stomoxys calcitrans) and lice (Bovicola bovis and Linognathus vituli)61 may play an important role in the transmission of infection amongst stabled animals.70 Eperythrozoon spp. can also be easily transmitted iatrogenically by all parenteral routes with infected blood or organ suspensions. Eperythrozoon suis is transmitted transplacentally.4 In utero infection of E. ovis has not been demonstrated in sheep.15
The role that wild animals may play as reservoir hosts would seem to be unimportant as it is believed that Eperythrozoon spp. are limited in their host ranges.24 Their host specificity is poorly defined and this is one aspect which has cast doubt on the taxonomic position of some of them.48 Neitz61 showed that E. ovis and E. wenyoni are both hostspecific in domestic stock. Hoyte37 confirmed this finding and also showed that E. tagenodes is not infective to sheep. However, E. ovis may be infective to goats,16 natural infections in goats with E. ovis having been reported in both South Africa95 and Australia.12 Latent infections have also been produced in two blesbuck (Damaliscus albifrons)60 and an eland (Taurotragus oryx) following artificial infection with E. ovis.18
Neitz59 reported that in sheep age makes no difference to the susceptibility to infection with E. ovis in South Africa. But in contrast, workers in some countries, especially Australia and New Zealand, describe various degrees of susceptibility among lambs, young sheep and adult sheep. Infection may result in subclinical infections, in an ill-thrift syndrome, or in severe clinical disease with high morbidity and mortality.8
No detailed studies have been made on eperythrozoonosis of sheep and goats in southern Africa. According to Neitz59 farmers in South Africa describe an anaemic condition in sheep to which the name ‘bleeksiekte’ (pallor disease) has been given. The aetiology of this condition remains obscure, and it is unknown to what extent verminosis and malnutrition or other contributory factors may be involved. The epidemiology of E. ovis infections in sheep is probably further complicated in endemic areas by other haemoparasitic diseases such as anaplasmosis and theileriosis.
Once animals are infected they remain life-long carriers. Sheep that recover from E. ovis infection develop a premunity; artificially infected sheep may be infected for up to ten and a half years.14 Autosterilization has not been reported, but chemosterilization renders sheep fully susceptible to reinfection. 62, 76 Splenectomy or treatment with dexamethasone results in relapses and a resurgence of parasitaemia.25, 51, 59 Lambs that suckle E. ovis-carrier ewes appear to be protected from patent infection.15 Serum antibody titres (which are believed to be protective) of uninfected lambs of such ewes apparently decline to zero within three weeks after weaning.15
One of the biggest obstacles faced when producing redwater (babesiosis) live-blood vaccine for cattle at Onderstepoort is the interference caused by concurrent E. wenyoni, E. tuomii and E. teganodes infections in splenectomized donor calves. These infections result in prolonged incubation periods and in the suppression of parasitaemias caused by both Babesia bovis and B. bigemina.70 Interference between concurrent eperythrozoon and other haemoparasitic infections, such as between E. wenyoni and Anaplasma marginale20 and between E. ovis and Babesia ovis,23 have also been reported. Eperythrozoon wenyoni infection not only causes extension of the incubation period in B. bigemina infection,35 but it also suppresses the parasitaemia of A. marginale and B. bigemina infections.57 Eperythrozoon teganodes apparently suppressed the development of A. marginale in a splenectomized calf.27 Complement fixation testing has shown that Anaplasma and Eperythrozoon spp. share antigens.46 However, the indirect fluorescent antibody test shows no cross-reactivity between E. ovis antigen and Anaplasma ovis antisera.40 Even between members of the Eperythrozoon spp., such as between E. tuomii and E. wenyoni, there appears to be no antigenic relationship.97
Eperythrozoon suis and E. parvum infections result in a subclinical carrier state which may be activated under stress. There is no cross-immunity between these two species, 84 but infection of E. suis suppresses the development of parasitaemia of E. parvum.85 Eperythrozoon parvum also inhibits the development of parasitaemia of Babesia trautmanni infection.3
Pathogenesis
Little information is available on the pathogenesis of Eperythrozoon spp. infections.
The majority of Eperythrozoon spp. infections, including those due to more pathogenic species, are subclinical and may be exacerbated by stress. Diseases produced by Eperythrozoon spp. are characterized by the destruction of erythrocytes or, in the case of E. tuomii, thrombocytes.
It has been suggested that anaemia in ovine eperythrozoonosis is not directly associated with the parasitaemia, but that it could have what was called an allergic base.94 Eperythrozoon ovis also seems to affect glutathione, which is essential for normal erythrocyte integrity.89, 92 Heinz bodies in erythrocytes of E. ovis-infected sheep56, 67, 92 have also been implicated in the pathogenesis of anaemia.56
Certain aspects of E. ovis infections are consistent with those of a haemolytic anaemia.19, 59 While Sutton90, 91 believes that intravascular haemolysis occurs, Sheriff78 concludes that haemoglobinuria, as reported by Overäs,68 is rarely found in E. ovis infection, which supports the contention that extravascular erythrophagocytosis takes place.
The cause of anaemia in eperythrozoonosis in pigs has been ascribed to autoimmune haemolysis cause by ‘cold’ auto-antibodies.33 These antibodies are attached to erythrocytes and induce destruction by phagocytosis, primarily in the spleen, but also in lymph nodes.69 In vitro agglutination of sensitized erythrocytes occurs spontaneously at 25 and 4 °C respectively.103
The agglutination phenomenon correlates with increases of serum antibodies (as measured by the indirect haemagglutination test) and of total serum globulin concentrations associated with the appearance of IgM ‘cold’ agglutinins.104 Erythrocytes, lost to the ‘cold’ agglutinin phenomenon, are replaced by reticulocytes. As the new red blood cells are not readily infected and the altered erythrocytes are phagocytosed, the antigenic stimulus required for the maintenance of indirect haemagglutination titres and globulin concentrations is effectively removed. 103 Haematological findings in splenectomized pigs indicate the development of a normochromic anaemia caused by extravascular erythrolysis.28
Natural E. suis infections show no signs of penetration and erosion of the red blood cell membrane comparable to those which occur in Haemobartonella felis infections.69 Severe deformation of the red blood cell membrane has, however, been demonstrated in splenectomized pigs infected experimentally with E. suis.103 It is thought that the humoral immune response is directed in part against these parasitealtered erythrocytes.103 Autoantibodies to erythrocytes, possible mechanisms involved in parasitic immunoregulation, and misdirected immune responses have been implicated in E. suis infections.103 Smith, as cited by Henry,30 believes that E. suis in pigs may affect all age groups and that its clinical manifestation is not limited to the classic disease of anaemia and icterus of feeder pigs under stress.
The only information regarding the pathogenesis of infections by E. wenyoni, E. teganodes and E. tuomii has been obtained from studies using splenectomized animals.45, 98, 105 In none of these studies were significant ultrastructural changes of infected red blood cells or thrombocytes observed.45
Clinical signs
Neitz and his co-workers60, 62, 64 showed that the incubation period following artificial infection with E. ovis could be as short as 4 to 15 days, but could sometimes extend to several weeks. Parasitaemia appears concurrently with the rise in body temperature and increases until the first signs of anaemia are detected five to eight days after the initial detection of parasitaemia. Parasitaemia then wanes abruptly; few parasites are seen when the anaemia is most severe. Recovered animals remain carriers and recrudescent parasitaemias may occur at irregular intervals for six weeks or longer.
The severity of the disease in sheep is extremely variable and seems to depend on the degree of the parasitaemia and its duration.25 A continuous or intermittent febrile reaction may be followed by variable anaemia and sometimes icterus.63 Other signs include depression, anorexia, loss of condition, debility, retardation of growth rate, lowered haemoglobin levels, a drop in wool production and decreased exercise tolerance.11, 76 The pulse rate becomes rapid and weak and the respiration accelerates. Haemoglobinuria occurs very rarely.59 Mortality rate is usually very low.8, 51, 56
In pigs clinical signs are most likely to be observed in young suckling piglets and sows shortly after farrowing. Most E. suis infections result in a subclinical carrier state which may lead to overt disease when animals are stressed53 The early phase of the infection is characterized by high fever, but in more advanced cases temperatures may be normal or subnormal, and the mucous membranes may be pale and icteric. Other signs include weakness, depression, loss of appetite, constipation, bile-stained faeces and dyspnoea.47 Some animals suffering from the disease may die. In animals that recover relapses may occur.
Eperythrozoon wenyoni usually causes only mild fever and anaemia in intact cattle65 but very occasionally some affected animals manifest severe clinical signs.71, 73 Severe anaemia and icterus occur in infected splenectomized animals.61, 93
Hoyte36 considered E. teganodes to be of similar pathogenicity to E. wenyoni. Eperythrozoon tuomii have been reported to cause fever and anaemia in splenectomized cattle.96, 99, 100, 105
Pathology
The major lesions caused by eperythrozoa (except E. tuomii) are the consequence of erythrocyte destruction resulting from erythrophagocytosis or intravascular haemolysis and the stimulation of the immune system, which leads to hyperplasia of the reticuloendothelial cells.
In sheep, lowered haematocrit and haemoglobulin levels, hypochromic anaemia, a rise in leukocyte counts and erythrophagocytosis have been reported.59, 63 In some animals, lowered haemoglobin and haematocrit levels occur one week after peak parasitaemia,10 whereas in others, there is a relationship between the severity of anaemia and peak parasitaemia.55, 67 The main chemical pathological features include an increase in blood lactic acid and a lowered blood pH.87, 88 Pyruvate and lactate concentrations reduce the blood pH 10 to 14 days after infection.42 Sutton87, 88 reported that affected erythrocytes have a high glycolytic activity. On the other hand, Ilemobade and Blotkamp42 found no effect on the concentration of blood glucose in infected animals.
Variable degrees of anaemia, icterus, and fatty degeneration of the liver have been reported in sheep.2, 8, 19, 44, 51, 59,67, 90 The spleen is enlarged and shows a reddish-brown pulp and severe hyperplasia of the Malpighian bodies. There may also be a marked hydropericardium together with subcutaneous oedema along the ventral aspect of the neck. Haemosiderin is present in the kidneys, liver and spleen at peak and late stages of parasitaemia.
Necropsy findings in pigs include anaemia, icterus, a yellowish- brown enlarged liver, and splenomegaly. There may also be hydropericardium, ascites, and epicardial haemorrhages. Microscopic lesions include hyperplasia of the red bone marrow, haemosiderosis in the spleen and liver, and fatty changes and necrosis of hepatocytes.17, 32, 38, 47 Perivascular oedema and discrete glial mobilization in the brain have been described in E. suis carrier pigs that had been splenectomized.9
Eperythrozoon wenyoni infections in splenectomized calves result in lowered packed red blood cell volumes, a rise in white blood cell counts and erythrophagocytosis.61, 71 Changes in splenectomized calves infected with E. wenyoni include an increase in lactic acid, lowered blood pH, negative base excess and lowered standard bicarbonate content.106 Metabolic acidosis has also been reported.101
Eperythrozoon tuomii infection results in a prolonged blood-clotting time and severe reduction in the number of thrombocytes and petechiae in the organs of splenectomized calves.105
Diagnosis
Eperythrozoon spp. infections should be considered in anaemic animals, especially if they have recently been splenectomized or immunosuppressed. In most cases, eperythrozoonosis cannot be diagnosed clinically until anaemia is well advanced. If the examination of blood smears fails to confirm the diagnosis, subinoculation of blood into susceptible splenectomized hosts may be considered.
Successful diagnosis of eperythrozoonosis is often dependent upon the correct technique being used when making a blood smear. Dirty glass slides and stain deposits can hamper recognition of these delicate parasites. Blood smears should be made from fresh blood preferably collected directly from the live animal, as the organisms tend to lose their structural integrity and may not be identifiable in smears made from stored blood.7 For example E. wenyoni ‘disappeared’ from the blood of a splenectomized calf within four hours of death.70 If stored blood is used, blood collected in EDTA is better for diagnostic purposes than heparinized or citrated blood.8 It has also been demonstrated that E. ovis is frequently absent in smears made from the peripheral blood of sheep in which anaemia is advanced.8, 51, 59, 79
The thick blood smear technique54 for Babesia spp. identification is extremely useful for making early diagnoses of eperythrozoonosis.70 Different scoring systems are used to quantify the parasitaemias.7
The organisms appear bright orange when stained with acridine orange and viewed under a fluorescent microscope. 24 This staining technique is more reliable for demonstrating low infections than is the Giemsa stain.25
Campbell et al.8 believe that E. ovis infections result in a characteristic type of anaemia, apparently of diagnostic value, which is predominantly either macro- or normocytic and normochromic in nature. Hoffmann31 contends that clinical signs in pigs are non-specific and that the phenomenon of instant agglutination of blood samples is important in the diagnosis of E. suis.
A variety of serological techniques can be used to diagnose infection on a flock or herd basis. These tests include direct immunofluorescence,46 the modified antiglobulin (Coombs) test,80 indirect haemagglutination,82, 83 complement fixation,10 indirect immunofluorescence40, 41, 64, 66 and enzyme-linked immunosorbent assay (ELISA).49
Differential diagnosis
Any condition that results in anaemia and icterus in sheep, goats, cattle and pigs may be confused with eperythrozoonosis. In sheep these include malnutrition, helminthosis, and the syndrome known as ill-thrift (Ill thrift). Differentiation between anaemia caused by copper and cobalt imbalances and E. ovis infections is difficult.8, 76 Recent work in Australia has shown that E. ovis is the major cause of icterus in lambs and that copper poisoning is much less involved.2
The macroscopic lesions caused by E. ovis in sheep closely resemble those seen in ovine anaplasmosis60 or chronic copper poisoning.2
In South Africa, concurrent Anaplasma ovis and Theileria ovis (T. separata?) infections may aggravate E. ovis infections in sheep.
The poisoning of goats with the pods of Acacia nilotica braussiana may result in the development of anaemia, icterus, methaemoglobinaemia and haemoglobinaemia.95
Babesia trautmanni infections and leptospirosis should be differentiated from E. suis infections in pigs.29
Control
Different drugs have been tested for activity against eperythrozoonosis. 24 Arsenicals and tetracyclines appear to be effective against most infections,27, 99, 100, 105 but in spite of their efficacy, these drugs do not sterilize the infection in most cases.7, 62, 72 Treatment of sheep infected with E. ovis with spirotrypan sterilizes the infection, and such animals become fully susceptible to reinfection.77
The arsenical drug, neosalvarsan, is effective in the treatment of E. ovis and E. wenyoni infections.61, 63
Both arsenicals, which are becoming increasingly difficult to obtain commercially, and oxytetracyclines at a dosage rate of 10 mg/kg body weight administered intravenously or intramuscularly, are regularly and successfully used to treat E. wenyoni, E. tuomii, E. teganodes and E. ovis infections in splenectomized cattle and sheep at the Onderstepoort Veterinary Institute.70 In acute cases it may be necessary to repeat the treatment if the febrile reaction and/or parasitaemia persists, and during parasitic relapses. According to Campbell et al.,8 treatment of affected sheep should be based on the reduction of stress by good husbandry rather than by therapy.
Tetracyclines appear to be the drugs of choice for treating E. suis infections.28, 30, 38, 58, 74 Eperythrozoon suis infections do not seem to have an appreciable effect on reproductive performance,104 but chlortetracycline treatment in the diet of sero-positive sows results in increased productivity.74
Although not conclusively proven, the possible role of ectoparasites in the transmission of the infections should be considered. It is therefore recommended that animals be treated for ectoparasites during outbreaks. Care should be exercised to prevent iatrogenic transmission of infection.
References
- ADLER, S. & ELLENBOGEN, V., 1934. A note on two new blood parasites of cattle, Eperythrozoon and Bartonella. Journal of Comparative Pathology and Therapeutics, 47, 219–221.
- ARCHER, J.F. & LITTLEJOHNS, I.R., 1984. Eperythrozoon ovis and copper poisoning as causes of jaundice in lamb carcases. Australian Veterinary Journal, 61, 312–313.
- BARNETT, S.F., 1963. Eperythrozoonosis in pigs in Kenya. Bulletin of Epizootic Diseases of Africa, 2, 185–195.
- BERRIER, H.H. & GOUGE, R.E., 1954. Eperythrozoonosis transmitted in utero from carrier sows to their pigs. Journal of the American Veterinary Medical Association, 124, 98–100.
- BRUYNOGHE, R. & VASSILIADIS, P., 1929. Contribution a l’étude des eperythrozoaires coccoides. Annales de Parasitologie Humaine et Compare’e, VII, 353–363.
- CAGNASSO, A., ABATE, O., GUGLIELMINO, R. & DOTTA, U., 1983. Segnalazione di Eperythrozoonosi in vitelli. Schweizer Archiv für Tierheilkunde, 125, 353–357.
- CALLOW, L.L., 1984. Protozoal and Rickettsial Diseases. Vol. 5. Canberra: Australian Bureau of Animal Health. Australian Government Publishing Service.
- CAMPBELL, R.W., SLOAN, C.A. & HARBUTT, P.R., 1971. Observations on mortality in lambs in Victoria associated with Eperythrozoon ovis. Australian Veterinary Journal, 47, 538–541.
- CRUZ, E., DELGADO, L., LOPEZ, R., POLANCO, H., LOMBILLO, R.D. & RODRIQUEZ, O.N., 1985. Organ pathology in experimental porcine eperythrozoonosis. Revista de Salud Animal, 7, 59–66.
- DADDOW, K.N., 1977. A complement fixation test for the detection of Eperythrozoon infection in sheep. Australian Veterinary Journal, 53, 139–143.
- DADDOW, K.N., 1979. Eperythrozoon ovis—a cause of anaemia, reduced production and decreased exercise tolerance in sheep. Australian Veterinary Journal, 55, 433–434.
- DADDOW, K.N., 1979. The natural occurrence in a goat of an organism resembling Eperythrozoon ovis. Australian Veterinary Journal, 55, 605–606.
- DADDOW, K.N., 1980. Culex annulirostris as a vector of Eperythrozoon ovis infection in sheep. Veterinary Parasitology, 7, 313–317.
- DADDOW, K.N., 1981. The duration of the carrier state of Eperythrozoon ovis infection in sheep. Australian Veterinary Journal, 57, 49.
- DADDOW, K.N., 1982. The protection of lambs from eperythrozoon infection while suckling Eperythrozoon ovis carrier ewes. Veterinary Parasitology, 10, 41–45.
- DONATIEN, A. & LESTOQUARD, F., 1935. Existence d’Eperythrozoon ovis en Algérie. Bulletin de la Société de Pathologie Exotique, 28, 423–426.
- DUNLAP, J.S., 1975. Protozoa. In: dunne, h.w. & leman, a.d., (eds). Diseases of Swine. 4th edn. Ames, Iowa: Iowa State University Press.
- ENIGK, K., 1942. Die Emphäglichkeit der Elenantilope für Anaplasma ovis und Eperythrozoon ovis. Deutsche Tropenmedizinische Zeitschrift, 46,
- FOGGIE, A. & NISBET, D.I., 1964. Studies on Eperythrozoon infection in sheep. Journal of Comparative Pathology, 74, 45–61.
- FOOTE, L.E., LEVY, H.E., TORBERT, B.J. & OGLESBY, W.T., 1957. Interference between anaplasmosis and eperythrozoonosis in splenectomized cattle. American Journal of Veterinary Research, 18, 556–559. 48–52.
- FOURIE, N. & PROZESKY, L., 1989. Veterinary Research Institute, Onderstepoort. Unpublished data.
- FRIEDHOFF, K.T., 1982. Rickettsieninfektionen (Ehrlichia, Eperythrozoon, Haemobartonella) bei Haustieren in Deutschland. Fortschritte der Veterinärmedicin, supplement to Zentralblatt für Veterinärmedizin, 35, 204–209.
- FRIEDHOFF, K., DROMMER, W. & WOLFHAGEN, M., 1971. Infektionen mit Eperythrozoon ovis bei Schafen in Norddeutschland. Berliner und Münchener Tierärtzliche Wochenschrift, 84, 361–380.
- GOTHE, R. & KREIER, J.P., 1977. Aegyptianella, Eperythrozoon and Haemobartonella. In: kreier, j.p., (ed.). Parasitic Protozoa. New York, San Francisco and London: Academic Press.
- GULLAND, F.M., DOXEY, D.L. & SCOTT, G.R., 1987. Changing morphology of Eperythrozoon ovis. Research in Veterinary Science, 43, 88–91.
- HADANI, A., DE HAAN, L., GUGLIELMONE, A.A. & ANZIANI, O.S., 1980. Eperythrozoonosis en terneros en la Argentina. Revista de Medicina Veterinaria, 61, 9–19.
- HADANI, A., GUGLIELMONE, A.A., ANZIANI, O.S., TARABLAS, H., MANGOLD, A., BERNUCEZ, A., CASTOR, J.C. & GONZALEZ DE RIOS, L., 1982. A case of apparent suppression of Anaplasma marginale infection by eperythrozoonosis (Eperythrozoon teganodes). Veterinary Parasitology, 9, 267–287.
- HEINRITZI, K., WENTZ, I. & BOLLWAHN, W., 1984. Haematologische Befunde bei der akuten Eperythrozoonose der Schweine. Berliner und Münchener Tierärtzliche Wochenschrift, 97, 404–407.
- HENNING, M.W., 1956. Animal Diseases in South Africa. Pretoria, South Africa: Central News Agency, Ltd.
- HENRY, S.C., 1979. Clinical observations on eperythrozoonosis. Journal of the American Veterinary Medical Association, 174, 601–603.
- HOFFMANN, H., 1984. Symptomatologie und Diagnostik der Eperythrozoonose in Ferkelerzeugerbetrieben. Tierärtzliche Umschau, 39, 474–479.
- HOFFMANN, R. & SAALFELD, K., 1977. Ausbruch einer Eperythrozoonose in einem Schweinenastbestand. Deutsche Tierärtzliche Wochenschrift, 84, 7–9.
- HOFFMANN, R., SCHMID, D.O. & HOFFMANN-FEZER, G., 1981. Erythrocyte antibodies on porcine eperythrozoonosis. Veterinary Immunology and Immunopathology, 2, 111–119.
- HOWARD, G.W., 1975. The experimental transmission of Eperythrozoon ovis by mosquitoes. Parasitology, 71, xxxiii.
- HOYTE, H.M.D., 1961. Initial development of infections with Babesia bigemina. Journal of Protozoology, 8, 462–466.
- HOYTE, H.M.D., 1962. Eperythrozoon teganodes sp. nov. (Rickettsiales), parasitic in cattle. Parasitology, 52, 526–532.
- HOYTE, H.M.D., 1971. The infectivity of Theileria mutans, Eperythrozoon wenyoni and E. teganodes to sheep and of Eperythrozoon ovis to cattle. British Veterinary Journal, 127, 1–3.
- HSU, F.S., CHUNG, W.B., HU, D.G., CHOU, N.Y. & SHIEH, W.I., 1985. Outbreak and treatment of eperythrozoonosis in swine in Taiwan. Journal of the Chinese Society of Veterinary Science, 11, 211–220.
- ICHIJO, S., HOSOKAWA, S., KIM, D. & KONISHI, T., 1982. Scanning and transmission electron microscopic observation of Eperythrozoon ovis. Japanese Journal of Veterinary Science, 44, 127–132.
- ILEMOBADE, A.A. & BLOTKAMP, C., 1978. Eperythrozoon ovis. I. Serological diagnosis of infection by the indirect immunofluorescent antibody test. Tropenmedizin und Parasitologie, 29, 307–310.
- ILEMOBADE, A.A. & BLOTKAMP, C., 1978. Eperythrozoon ovis. II. Prevalence studies in sheep in Nigeria using the indirect immunofluorescent antibody test. Tropenmedizin und Parasitologie, 29, 311–314.
- ILEMOBADE, A.A. & BLOTKAMP, C., 1978. Eperythrozoon ovis. III. The effect of infection on blood pH, concentrations of pyruvate, lactate and glucose in blood, haematology, and on concurrent and superimposed Trypanosoma vivax infection in sheep. Tropenmedizin und Parasitologie, 29, 443–450.
- JANSEN, B.C., 1952. The occurrence of Eperythrozoon parvum Splitter, 1950 in South African swine. Onderstepoort Journal of Veterinary Research, 25, 5–6.
- JOLLY, R.D., 1967. Eperythrozoon ovis infection in a lamb. New Zealand Veterinary Journal, 15, 47–48.
- KEETON, K.S. & JAIN, N.C., 1973. Eperythrozoon wenyoni: A scanning electron microscope study. Journal of Parasitology, 59, 867–873.
- KREIER, J.P. & RISTIC, M., 1963. Morphologic, antigenic and pathogenic characteristics of Eperythrozoon ovis and Eperythrozoon wenyoni. American Journal of Veterinary Research, 24, 488–500.
- KREIER, J.P. & RISTIC, M., 1968. Haemobartonellosis, Eperythrozoonosis, Grahamellosis and Erhlichiosis. In: weinman, d. & ristic, m., (eds). Infectious Blood Diseases of Man and Animals. New York, London: Academic Press.
- KRIEG, N.R. & HOLT, J.G., 1984. Bergey’s Manual of Systematic Bacteriology. Baltimore: Williams and Wilkins.
- LANG, F.M., FERRIER, G.R. & NICHOLLS, T.J., 1987. Detection of antibodies to Eperythrozoon ovis by the use of an enzyme-linked immunosorbent assay. Research in Veterinary Science, 41, 249–252.
- LEEFLANG, P., 1978. Prevalence and significance of tick-borne diseases of domestic stock in northern Nigeria. In: wilde, j.k.h., (ed.). Tick-borne Diseases and their Vectors. Edinburgh: University Press.
- LITTLEJOHNS, I.R., 1960. Eperythrozoonosis in sheep. Australian Veterinary Journal, 36, 260–265.
- MACKEE, A.E., ZIEGLER, R.F. & GILES, R.C., 1973. Scanning and transmission electron microscopy of Haemobartonella canis and Eperythrozoon ovis. American Journal of Veterinary Science, 34, 1196–1201.
- MADSEN, M., 1986. Eperythrozoonose hos svin—En overset sygdom? Nordisk Veterinaermedicin, 38, 57–67.
- MAHONEY, D.F. & SAAL, J.R., 1961. Bovine babesiosis: Thick blood films for the detection of parasitaemia. Australian Veterinary Journal, 37, 44–47.
- MARTIN, B.J., CHRISP, C.E., AVERILL, D.R. & RINGLER, D.A., 1986. Eperythrozoon ovis, a health risk for research sheep (Abstract). Laboratory Animal Science, 36, 564.
- MASON, R.W., MANUEL, M., DADDOW, K.N. & WITT, D.J., 1980. Eperythrozoon ovis recorded in Tasmania in association with Heinz body development. Australian Veterinary Journal, 57, 46.
- MCHARDY, N., 1974. Elimination of Eperythrozoon spp. from mixed infections with Babesia and Anaplasma. International Journal for Parasitology, 4, 107–108.
- MULLER, E. & NEDDENRIEP, G., 1979. Eperythrozoon in einen Ferkelerzeugerbetrieb in Norddeutschland. Praktische Tierärzt, 60, 662–665.
- NEITZ, W.O., 1937. Eperythrozoonosis in sheep. Onderstepoort Journal of Veterinary Science and Animal Industry, 9, 9–30.
- NEITZ, W.O., 1939. The immunity in heartwater. Onderstepoort Journal of Veterinary Science and Animal Industry, 13, 245–283.
- NEITZ, W.O., 1940. Eperythrozoonosis in cattle. Onderstepoort Journal of Veterinary Science and Animal Industry, 14, 9–28.
- NEITZ, W.O., 1968. Eperythrozoon ovis infection. Bulletin Office International des E’pizooties, 70, 373–378.
- NEITZ, W.O., ALEXANDER, R.A. & DU TOIT, P.J., 1934. Eperythrozoon ovis (sp. nov.) infection in sheep. Onderstepoort Journal of Veterinary Science and Animal Industry, 3, 263–271.
- NICHOLLS, T.J. & VEALE, P.I., 1986. A modified indirect immunofluorescent assay for the detection of antibody to Eperythrozoon ovis in sheep. Australian Veterinary Journal, 63, 157–159.
- OBI, T.U. & ANOSA, V.O., 1980. Haematological studies on domestic animals in Nigeria. IV. Clinico-haematological features of bovine trypanosomiasis, theileriosis, anaplasmosis, eperythrozoonosis and helminthiasis. Zentralblatt für Veterinärmedizin, 27, 789–797.
- ODHER, H., 1967. Some observations on Eperythrozoon infection in non-splenectomized sheep and the detection of the parasites and their antibodies by immunofluorescence. Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene, Erste Abteilung Orginale, A 203, 391–401.
- OVERÄS, J., 1969. Studies on Eperythrozoon ovis infection in sheep. Acta Veterinaria Scandinavica, Supplement, 28, 1–148.
- OVERÄS, J., 1987. Haemoglobinuria in eperythrozoonosis in sheep. The Veterinary Record, 121, 277.
- POSPISCHILL, A. & HOFFMANN, R., 1982. Eperythrozoon suis in naturally infected pigs: A light and electron microscopic study. Veterinary Pathology, 19, 651–657.
- POTGIETER, F.T., 1989. Veterinary Research Institute, Onderstepoort. Unpublished data.
- PURNELL, R.E., BROCKLESBY, E.W. & YOUNG, E.R., 1976. Eperythrozoon wenyoni, a possible cause of anaemia in British cattle. The Veterinary Record, 98, 411.
- PURNELL, R.E., LEWIS, D., HOLMAN, M.R. & YOUNG, E.R., 1981. Investigations on a Babesia isolated from Scottish sheep. Parasitology, 83, 347–356.
- QUINLAN, J.F., 1985. Suspected eperythrozoonosis in dairy cows. Irish Veterinary Journal, 39, 2–27.
- ROSENKRANS, C.J., ZINN, G.M. & JESSE, G.W., 1984. Chlortetracycline as a treatment against the effect of eperythrozoonosis on sow productivity. Proceedings of the Eighth International Pig Veterinary Society Congress, 27–31 August 1984, Ghent, Belgium.
- SCHILLING, V., 1928. Eperythrozoon coccoides eine neue durch Splenectomie aktivierbare Dauerinfektion der weisse Maus. Klinische Wochenschrift, 7, 1854–1855.
- SHERIFF, D., 1972. Diagnosis of Eperythrozoon ovis infection. Australian Veterinary Journal, 48, 128.
- SHERIFF, D., 1973. The effect of spirotrypan on Eperythrozoon ovis in sheep. The Veterinary Record, 93, 288–289.
- SHERIFF, D., 1978. The pathology of Eperythrozoon ovis. New Zealand Veterinary Journal, 26, 315.
- SHERIFF, D., CLAPP, K.H. & REID, M.A., 1966. Eperythrozoon ovis infection in South Australia. Australian Veterinary Journal, 42, 169–176.
- SHERIFF, D. & GEERING, M.C., 1969. The antiglobulin (Coombs) test in Eperythrozoon ovis infection in sheep. Australian Veterinary Journal, 45, 505–507.
- SIMPSON, C.F. & LOVE, J.N., 1970. Fine structure of Haemobartonella bovis in blood and liver of splenectomized calves. American Journal of Veterinary Research, 31, 225–231.
- SISK, D.B., COLE, J.R. & PURNELL, A.R., 1980. Serologic incidence of eperythrozoonosis in Georgia swine. 23rd Annual Proceedings, College of Veterinary Medicine, University of Georgia.
- SMITH, A.R. & RAHN, R., 1975. An indirect hemagglutination test for the diagnosis of Eperythrozoon suis infection in swine. American Journal of Veterinary Research, 36, 1319–1321.
- SPLITTER, E.J., 1950. Eperythrozoon suis n.sp. and Eperythrozoon parvum n.sp., two new blood parasites of swine. Science, III, 513–514.
- SPLITTER, E.J., 1953. Eperythrozoon parvum a filterable blood parasite of swine. Nature, 172, 40.
- SPLITTER, E.J. & WILLIAMSON, R.L., 1950. Eperythrozoonosis in swine. Journal of the American Veterinary Medical Association, 116, 360–364.
- SUTTON, R.H., 1976. Eperythrozoon ovis: The difference in carbohydrate metabolism between infected and uninfected sheep erythrocytes. Ajebak, 54, 449–458.
- SUTTON, R.H., 1977. The effect of Eperythrozoon ovis infection on the glucose level and some acid-base factors in the venous blood of sheep. Australian Veterinary Journal, 53, 478–481.
- SUTTON, R.H., 1978. Eperythrozoon ovis: The difference in carbohydrate metabolism between infected and uninfected sheep erythrocytes. Australian Journal of Experimental Biology and Medical Science, 54, 449–458.
- SUTTON, R.H., 1978. Observation on the pathology of Eperythrozoon ovis infection in sheep. New Zealand Veterinary Journal, 26, 229–230.
- SUTTON, R.H., 1978. The pathology of Eperythrozoon ovis. New Zealand Veterinary Journal, 27, 18.
- SUTTON, R.H., 1979. Effect of Eperythrozoon ovis and the reductive potential of sheep erythrocytes. Veterinary Parasitology, 5, 11–15.
- SUTTON, R.H., CHARLESTON, W.A.G. & COLLINS, G.H., 1977. Eperythrozoon wenyoni—a blood parasite of cattle. New Zealand Veterinary Journal, 25, 8–9.
- SUTTON, R.H. & JOLLY, R.D., 1973. Experimental Eperythrozoon ovis infection of sheep. New Zealand Veterinary Journal, 21, 160–166.
- TERBLANCE, M., PIENAAR, J.G., BIGALKE, R. & VAHRMEYER, J., 1967. Acacia nilotica (L.) Del. subsp. kraussiana (Benth.) Brenan. as a poisonous plant in South Africa. Journal of the South African Veterinary Medical Association, 38, 57–63.
- TUOMI, J., 1966. A micro-organism affecting bovine platelets. Experientia, 22, 458–459.
- TUOMI, J. & TANSKANEN, R., 1980. Antigenic non-relationship of two bovine eperythrozoa demonstrated by the immunofluorescent method. Acta Veterinaria Scandinavica, 21, 699–701.
- TUOMI, J. & VON BONSDORF, C.H., 1967. Ultrastructure of a micro-organism associated with bovine platelets. Experientia, 23, 1–4.
- UILENBERG, G., 1965. Notes sur les Eperythrozoon de bovins a Madagascar. Revue d’Elevage et de Médecine Vétérinaire des Pays Tropicaux,18, 73–81.
- UILENBERG, G., 1967. Eperythrozoon tuomii, n.sp. (Rickettsiales), troisième espèce d’Eperythrozoon des bovins Madagascar. Revue d’Elevage et de Médecine Vétérinaire des Pays Tropicaux, 20, 563–569.
- WENSING, T.H., NOUWENS, G., SCHOTMAN, A.J.H., VERNOOY, J. & ZWART, D., 1974. The effect of Eperythrozoon wenyoni on the glucose level and acid-base balance of bovine blood in vivo and in vitro. Tijdschrift Diergeneeskunde, 99, 136–142.
- ZACHARY, J.F. & BASGALL, E.J., 1985. Erythrocyte membrane alterations associated with the attachment and replication of Eperythrozoon suis: A light and electron microscopic study. Veterinary Pathology, 22, 164–170.
- ZACHARY, J.F. & SMITH, A.R., 1985. Experimental porcine eperythrozoonosis: T-lymphocyte suppression and misdirected immune responses. American Journal of Veterinary Research, 46, 821–830.
- ZINN, G.M., JESSE, G.W. & DOBSON, A.W., 1983. Effect of eperythrozoonosis on sow productivity. Journal of the American Veterinary Medical Association, 182, 369–371.
- ZWART, D., LEEFLANG, P. & VAN VORSTENBOSCH, C.J.A.H.V., 1969. Studies on an Eperythrozoon associated with bovine thrombocytes. Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene, Erste Abteilung Originale, A 210, 83–105.
- ZWART, D., SCHOTMAN, A.J.H. & STRAVER, A.J.M., 1970. Metabolic changes during Eperythrozoon infection in cattle. Research in Veterinary Science, 11, 105–111.




