- Infectious Diseases of Livestock
- Part 3
- Paratuberculosis
- GENERAL INTRODUCTION: SPIROCHAETES
- Swine dysentery
- Borrelia theileri infection
- Borrelia suilla infection
- Lyme disease in livestock
- Leptospirosis
- GENERAL INTRODUCTION: AEROBIC ⁄ MICRO-AEROPHILIC, MOTILE, HELICAL ⁄ VIBROID GRAM-NEGATIVE BACTERIA
- Genital campylobacteriosis in cattle
- Proliferative enteropathies of pigs
- Campylobacter jejuni infection
- GENERAL INTRODUCTION: GRAM-NEGATIVE AEROBIC OR CAPNOPHILIC RODS AND COCCI
- Moraxella spp. infections
- Bordetella bronchiseptica infections
- Pseudomonas spp. infections
- Glanders
- Melioidosis
- Brucella spp. infections
- Bovine brucellosis
- Brucella ovis infection
- Brucella melitensis infection
- Brucella suis infection
- Brucella infections in terrestrial wildlife
- GENERAL INTRODUCTION: FACULTATIVELY ANAEROBIC GRAM NEGATIVE RODS
- Klebsiella spp. infections
- Escherichia coli infections
- Salmonella spp. infections
- Bovine salmonellosis
- Ovine and caprine salmonellosis
- Porcine salmonellosis
- Equine salmonellosis
- Yersinia spp. infections
- Haemophilus and Histophilus spp. infections
- Haemophilus parasuis infection
- Histophilus somni disease complex in cattle
- Actinobacillus spp. infections
- Actinobacillus equuli infections
- Gram-negative pleomorphic infections: Actinobacillus seminis, Histophilus ovis and Histophilus somni
- Porcine pleuropneumonia
- Actinobacillus suis infections
- Pasteurella and Mannheimia spp. infections
- Pneumonic mannheimiosis and pasteurellosis of cattle
- Haemorrhagic septicaemia
- Pasteurellosis in sheep and goats
- Porcine pasteurellosis
- Progressive atrophic rhinitis
- GENERAL INTRODUCTION: ANAEROBIC GRAM-NEGATIVE, IRREGULAR RODS
- Fusobacterium necrophorum, Dichelobacter (Bacteroides) nodosus and Bacteroides spp. infections
- GENERAL INTRODUCTION: GRAM-POSITIVE COCCI
- Staphylococcus spp. infections
- Staphylococcus aureus infections
- Exudative epidermitis
- Other Staphylococcus spp. infections
- Streptococcus spp. infections
- Strangles
- Streptococcus suis infections
- Streptococcus porcinus infections
- Other Streptococcus spp. infections
- GENERAL INTRODUCTION: ENDOSPORE-FORMING GRAM-POSITIVE RODS AND COCCI
- Anthrax
- Clostridium perfringens group infections
- Clostridium perfringens type A infections
- Clostridium perfringens type B infections
- Clostridium perfringens type C infections
- Clostridium perfringens type D infections
- Malignant oedema⁄gas gangrene group of Clostridium spp.
- Clostridium chauvoei infections
- Clostridium novyi infections
- Clostridium septicum infections
- Other clostridial infections
- Tetanus
- Botulism
- GENERAL INTRODUCTION: REGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Listeriosis
- Erysipelothrix rhusiopathiae infections
- GENERAL INTRODUCTION: IRREGULAR, NON-SPORING, GRAM-POSITIVE RODS
- Corynebacterium pseudotuberculosis infections
- Corynebacterium renale group infections
- Bolo disease
- Actinomyces bovis infections
- Trueperella pyogenes infections
- Actinobaculum suis infections
- Actinomyces hyovaginalis infections
- GENERAL INTRODUCTION: MYCOBACTERIA
- Tuberculosis
- Paratuberculosis
- GENERAL INTRODUCTION: ACTINOMYCETES
- Nocardiosis
- Rhodococcus equi infections
- Dermatophilosis
- GENERAL INTRODUCTION: MOLLICUTES
- Contagious bovine pleuropneumonia
- Contagious caprine pleuropneumonia
- Mycoplasmal pneumonia of pigs
- Mycoplasmal polyserositis and arthritis of pigs
- Mycoplasmal arthritis of pigs
- Bovine genital mycoplasmosis
- Neurotoxin-producing group of Clostridium spp.
- Contagious equine metritis
- Tyzzer's disease
- MYCOTIC AND ALGAL DISEASES: Mycoses
- MYCOTIC AND ALGAL DISEASES: Pneumocystosis
- MYCOTIC AND ALGAL DISEASES: Protothecosis and other algal diseases
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Epivag
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ulcerative balanoposthitis and vulvovaginitis of sheep
- DISEASE COMPLEXES / UNKNOWN AETIOLOGY: Ill thrift
- Eperythrozoonosis
- Bovine haemobartonellosis
Paratuberculosis
This content is distributed under the following licence: Attribution-NonCommercial CC BY-NC  View Creative Commons Licence details here
View Creative Commons Licence details here
NJ Maclachlan and M-L Penrith (Editors). C D Buergelt, M E Hines II and A L Michel, Paratuberculosis, 2019.
Paratuberculosis
Previous authors: C D BUERGELT, S S BASTIANELLO AND A L MICHEL
Current authors:
C D BUERGELT - Professor Emeritus, DVM, PhD, University of Florida, Florida, 32605, United States of America
M E HINES II - Professor Emeritus, Retired from University of Georgia, DVM, PhD, Diplomate American College of Veterinary Pathology, Pathology Department, University of Georgia, 939 Old Kincaid Road, Colbert, Georgia, 30628, United States of America
A L MICHEL - Professor: Bacteriology, DVM, Dr Med Vet, PhD, Faculty of Veterinary Science, University of Pretoria, Private Bag X04, Onderstepoort, Pretoria, Gauteng, 0110, South Africa
![]()
Introduction
Paratuberculosis or Johne’s disease is a chronic infectious disease of cattle, sheep, goats, camelids and certain captive or free-living wild ruminant species (bison, deer, elk, caribou). Sporadically, it occurs in several aberrant hosts such as equines, non-human primates, rodents, lagomorphs, canines, felines, birds, bears, and wild boars. The disease is caused by Mycobacterium avium subsp. paratuberculosis (MAP), a facultative intracellular acid-fast bacillus. In infected herds or flocks, the majority of animals are subclinically infected. Clinical signs usually develop in animals that are more than two years old and are manifested by progressive emaciation and/or chronic enteritis and diarrhoea.
Johne and Frothingham were the first to describe paratuberculosis in Germany in 1895 as an unusual case of tuberculosis in a cow.53 The true causative agent, M. avium subsp. paratuberculosis, was isolated and grown in pure culture in 1902.27 The disease was officially recognized as an infection distinct from tuberculosis in 1906, and it was suggested that it be named paratuberculosis.8
Paratuberculosis is distributed throughout the world, and has been reported from all continents. Prevalence of the disease is determined on herd and individual animal level. Prevalence rates vary for individual countries depending on the diagnostic tests used and management practices performed. Global herd level prevalence has been estimated to be between 30-50 per cent in dairy cattle.11 Significant economic losses result from reduced weight salvage at slaughter and lower milk production.
Aetiology
Mycobacterium avium subsp. paratuberculosis is genotypically related to Mycobacterium avium and is grouped together with the Mycobacterium avium-intracellulare (MAI) complex. It is a non-motile short Gram-positive strongly acid-fast rod measuring approximately 1–2 μm × 0,5 μm. In both faeces and tissues the organism occurs in the form of clumps, a characteristic that aids in its identification. A network of intercellular filaments is considered to be responsible for the clumping of the bacilli.19
Phenotypically distinct lineages of MAP have been recognized.103 Current classification of MAP strains has been achieved using different molecular techniques including genetic sequencing. Two major groups of strains are distinguished, namely “Sheep-type” or “Type S” and “Cattle-type” or “Type C”. The type C group is synonymous with the Type II strain. Type S strains comprise types I and III. The ovine pigmented sheep strain is mainly associated with subtype I. A different group of strains has been designated “Bison” or “B-type”. Analysis of B-type strains revealed that isolates from Indian bisons (Bos gaurus) are different from isolates from bisons in the USA (Bison bison). The Indian bison type is considered a sublineage of the type C strain. Isolates from goats mainly are C-type. Genetic heterogeneity and phenotypic differences exist between MAP strains, affecting growth, virulence, infection and geographic distribution.
Many techniques for the primary cultivation of MAP from faeces and tissues of infected animals have been developed (see Diagnosis).
Primary MAP colonies on slants are small, 1 to 5mm in diameter, moist, convex and usually white and unpigmented, but yellow-pigmented strains have, on occasion, been isolated from sheep.94 Counts of colonies on primary isolation can provide a rough assessment of the number of mycobacteria being shed. The two principal distinguishing cultural characteristics of MAP are its slow growth pattern and its dependence on exogenous mycobactin, which is used by the bacterium as an iron chelator for metabolic activity.61 Mycobacterium avium subsp. paratuberculosis requires higher levels of iron to sustain multiplication in vitro than do all the other Mycobacterium spp.76, 100 To be considered as MAP, the organism must primarily be acid-fast and mycobactin dependent.6
Mycobacterium avium subsp. paratuberculosis is relatively resistant to adverse environmental factors and can survive for at least 270 days in water, 264 days in bovine faeces, and 252 days in cattle slurry59 but is inactivated by most disinfectants.
Epidemiology
Cattle,121 sheep,25, 90 goats,113 and many species of captive artiodactylids36 and free-living ruminants, such as white-tailed deer (Odocoileus virginianus),46 elk (Cervus elaphus),64 bighorn sheep (Ovis canadensis),125 Rocky Mountain goats (Oreamnos americanus),125 and North American bison23 are highly susceptible to infection with MAP. Findings of MAP in aberrant, non-ruminant hosts mentioned in the Introduction raise the possibility that these species can function as reservoir hosts or passive shedders and play a role in the spread of the bacteria into the environment. With the exception of lagomorphs16, 92 and non-human primates43 other susceptible species usually do not exhibit clinical signs or pathologic changes as seen in natural hosts.
Animals are most susceptible to infection as neonates and cattle infected as adults are unlikely to develop fully-fledged clinical disease as they are usually culled from the herd for other reasons. Cattle infected as adults may, however, shed organisms into the environment and thus serve as vectors of the disease.
Several factors appear to contribute to the age-dependent susceptibility of young animals to infection with MAP. Suckling calves have a higher intestinal acidity compared to that of adult ruminants and this favours the survival of the organisms.84 Bacterial survival is furthermore enhanced by the presence of lactoferrin and transferrin that are present in the colostrum ingested by suckling ruminants and serve as a source of iron for the production of mycobactin by the mycobacteria. Maternal antibodies in the milk of infected animals, such as opsonin, may enhance the uptake of MAP by M-cells and enterocytes.71
Calves of domestic cattle (and probably the offspring of other susceptible species) may become infected in one of three ways:
- in utero — up to 20 per cent of infections are believed to be congenital,77, 95, 110
- ingestion of infected milk from cows harbouring the organism,109 or
- intake of MAP from e.g. faecally contaminated cows’ teats, pastures or other materials/objects in the environment.
Pass-through or faecal shedding of organisms ingested by an animal from a contaminated environment without the development of infection or disease in the animal concerned may occur.121 Mycobacterium avium subsp. paratuberculosis has been isolated from the semen of bulls,60 embryos91 and foetuses.77, 110
Stress, such as parturition, may convert a subclinical case into an overt clinical case. Other factors that may precipitate clinical disease are transport and a low plane of nutrition.27
There is a correlation between the prevalence of paratuberculosis and certain soil types,54, 57 with acid conditions (low pH) of soil or pasture enhancing spread of the disease, whereas alkaline (high pH) conditions inhibit its spread.57 The observation of geographic soil pH variations being correlated with high or low prevalence of paratuberculosis has led to the recommendation that lime be applied to soils in order to elevate their pH and hence reduce the spread of the disease.84 The effect of pH on the prevalence of paratuberculosis is apparently related to the effect of pH on iron uptake by MAP, where an acidic environment enhances iron uptake.
Paratuberculosis progresses through three stages of development. In stage I of the infection there is no appreciable faecal shedding of the organism or development of overt disease. The next stage, stage II, is the subclinical excretory phase of the disease during which the organism is shed into the environment. The terminal phase (stage III) is the clinical phase of the disease, which is characterized by intermittent or continuous shedding of the organisms, diarrhoea and weight loss.31, 121 Super-shedders in infected herds give rise to disproportionate numbers of MAP in the environment. Super-shedders may produce up to 10,000 times more MAP (>10,000 colony forming units/g faeces) and contribute to a two- to threefold increase in infectiousness. Most of these super-shedders in herds are asymptomatic.
The duration of each stage of infection depends on the age of the animal at the time of exposure and the number of organisms ingested.121 The majority of exposed animals in a given herd are subclinically infected, but may shed bacteria intermittently in their faeces. Clinical disease represents merely the tip of the iceberg of the total number of infected animals on the farm.
Pathogenesis
A comprehensive review on the pathogenesis of ruminant paratuberculosis was published by Clarke30 and others.44, 126
Most ruminants are infected orally as young animals but in utero infection also plays a significant role. Following ingestion of MAP the organisms pass through the upper gastrointestinal tract to the ileum where they are engulfed by microfold-epithelial cells (M-cells). These are specialized epithelial cells in the lymphoid patches or domes of the ileal mucosa that function as phagocytes and are thought to represent an important pathway for the direct transport of macromolecules and antigens to the intestinal lymphoid tissue. The organisms are subsequently transferred to dendritic (Langerhans) cells in the ileal lamina propria and macrophages in the submucosa.71 After the macrophages have phagocytosed the bacilli discharged from M-cells, they present them as antigens to lymphocytes belonging to the gut-associated lymphoid tissue (GALT), thus sensitizing the intestinal immune system to the presence of the bacteria. Transport via M-cells is a direct route of intestinal antigen delivery, which is effective in the initial phase of infection with MAP as it allows the bacilli to reach macrophages situated beneath the mucosal barrier. The predominant location of Peyer’s patches in the ileum may explain why intestinal lesions are usually located in this part of the intestinal tract. Infected macrophages may also migrate into intestinal lymphatics and hence spread the infection to mesenteric lymph nodes.
Invasion of villous enterocytes by MAP can also occur through transcellular and paracellular routes.10
The intracellular fate of MAP in macrophages is governed by various pathways that after phagosomal incorporation and phagolysosome fusion include replication in bacteriophorous vacuoles, antigen processing for partial degradation, and exocytotic pathway after complete degradation. Intracellular MAP has developed strategies to escape defense systems that include 1) escape from the phagosome into the cell cytoplasm, 2) avoidance of phagosomal maturation and phagosome-lysosome fusion, 3) modification of lysosome content, and 4) passive protection by the bacterial cell wall envelope. The ability to reduce apoptosis of infected macrophages is an alternative mechanism to evade host defense, as is interference with autophagy during infection to avoid restriction of replication.80
The bacteria do not produce toxins, but their multiplication in macrophages results in the release of both immune inhibitory and activating molecules. Structural glycolipids such as phosphoglycolipids and lipoarabinomannans associated with the bacillary cell wall are particularly effective as immunogens. Phosphoglycolipids can stimulate the production of B cells, act as scavengers for oxygen and nitrous oxygen radicals, and fix complement C3, which facilitates phagocytosis.129 Lipoarabinomannans are B cell immunogens and they inhibit lymphocyte proliferation, decrease gamma interferon activation, activity and antigen processing, and increase the production of tumour necrosis factor (TNF). The latter substance enhances the inflammatory reaction, aids in the development of the granulomatous lesions, and is also involved in the induction of clinical signs such as cachexia and pyrexia.106
Activated infected macrophages are involved in the induction of specific T cell responses inducing T helper cell, T suppressor cell, and cytotoxic T cell subpopulation proliferation.30 There are initially two subsets of T helper cells, TH1 and TH2.12
Once activated, these subsets produce different lymphokines that direct the propagation of either cell-mediated immunity or humoral immunity. TH1 cells produce predominantly interleukin (IL)-2, TNF and gamma interferon, which are all directed toward cell-mediated immunity, whilst TH2 cells produce predominantly IL-4, 5, 6 and 10, which are oriented towards humoral immunity. The manipulation of the two subsets of T helper cells is under the influence of IL-12.106
As for other mycobacterial diseases, such as leprosy and tuberculosis, paratuberculosis is considered a dynamic, spectral disease that fluctuates between periods of cell-mediated immunity when the host is successfully coping with the infective organisms to humoral immune responses when the host’s resistance to the organisms is lost. The clinical disease of paratuberculosis is a reflection of the immunological reaction of the host25 and consequently during the course of infection there are periods of remission and exacerbation of the disease.18, 25 Specific protective immunity occurs during the early stages of infection. Later, when organisms are released from lysed macrophages they stimulate a humoral response in which the IgG subclass of immunoglobulins predominate. Humoral immunity does not participate in the destruction of MAP but is, from a diagnostic viewpoint, important in the serological demonstration of infection. As an infected animal progresses towards the clinical phase of paratuberculosis, cell-mediated immunity is lost and animals become anergic. Anergy allows infected macrophages to enter the blood stream, resulting in bacteraemia and dissemination of the organism to distant organs, such as the liver, kidneys, lungs, reproductive tract and udder. The latter scenario can explain an in utero infection rate of up to 20 per cent in cattle.110
Experiments undertaken to investigate the early stages of the pathogenesis of the disease in cattle suggest that the organisms, besides being taken up by the ileal GALT, are also taken up by the tonsillar lymphoid tissue83, 93 and that one to four months later the infection becomes localized in specific mesenteric lymph nodes. After 6 to 14 months, the mesenteric and ileocaecal lymph nodes are consistently infected and some organisms may spread to the caecum, colon, liver and other tissues.83
The early histological lesions in calves one month after infection are difficult to identify. Small clusters of epithelioid cells and individual multinucleate Langerhans- type giant cells may be evident in the paracortical zones of intestinal lymph nodes and at the tips of intestinal villi, and small numbers of acid-fast bacilli may be found within these cells. After four months, lesions are more frequent and pronounced and consist of scattered groups of epithelioid cells and associated giant cells. After 14 months the granulomatous lesions are more extensive and numerous acid-fast bacteria are present in the giant cells and epithelioid cells. Epithelioid cells are derived from circulating monocytes of bone marrow origin while the Langerhans giant cells form from amitotic division of macrophages.22
The presence of severe granulomatous inflammation in the intestinal mucosa gives rise to the so-called villus-filling effect causing decreased assimilation and absorption of nutrients and exudation into the lumen of the intestinal tract with a resultant diarrhoea and wasting syndrome.22, 30 An antigen-antibody reaction with release of histamines and vasoactive amines also appears to be involved in the development of the diarrhoea.
As in leprosy in humans, distinct histomorphological forms of granulomatous responses may occur in bovine paratuberculosis.22 In some animals polyhedral or epithelioid macrophages with vacuolated cytoplasm that contain few to moderate numbers of acid-fast bacteria predominate, whilst in others, the macrophages are more histiocytic in type and contain numerous acid-fast bacilli. The significance of these different cell types is unknown.
Macrophages are the target cells in paratuberculosis. Once infection of macrophages has been accomplished the mycobacteria tend to evade and resist intracellular destruction by lysosomal and phagosomal enzymes and toxic reactive oxygen intermediates. Nitric oxide, however, apparently has no effect on intracellular growth of the bacteria. The destructive capabilities of macrophages are dependent on the presence of T cell and cytokine activity in their environment. There are limited studies on the effects of cytokines in paratuberculosis but in chronically infected cattle IL-1 activity of blood monocytes has been reported to be elevated.129 There is some information on the intestinal immune response in paratuberculosis. Phenotyping of intestinal T-cell lymphocytes in infected cattle indicated an increased presence of γδ + and CD8+ T-cell subsets rather than CD4+ cells12 and suggested that CD4+ mediated immunity is suppressed. Cytokine gene expression measurement in the intestine and mesenteric lymph nodes of infected cattle has suggested a role for gamma interferon in preventing progression of paratuberculosis from the asymptomatic to the clinical stage106 Expression of IL-4 did not differ between asymptomatic and clinical groups and progression of paratuberculosis to the clinical stage is associated with reduced expression of gamma interferon at the site of infection.106 Immunohistochemical staining of intestinal tissue of sheep with paratuberculosis demonstrated depleted concentrations of CD4+ and CD8+ cells, but no changes in the CD4+:CD8+ ratio.4 However, there was an increase in the ratio of γδ + to CD4+ and CD8+ cells.74 γδ + T-cells are typically located in the enteric mucosa and are also referred to as mucosal epithelial T-cells.62 In paratuberculous goats, the distribution of the various T cell phenotypes apparently depends on the severity of the lesions. In severely infected goats there was a decrease in the CD4+ T cell subpopulation and an increase of the CD8+ T cell in the intestines and mesenteric lymph nodes.74
A decreased number of CD4+ T cell lymphocytes is usually associated with the presence of major histocompatibility complex class II epithelioid macrophages containing numerous acid-fast bacilli. The data obtained from typing subsets of intestinal goat T cell lymphocytes suggested that paratuberculosis is a disease with many synergistic and antagonistic influences on the immune response, the balance of which determines the outcome of disease.111
Clinical signs
Clinical signs are usually seen in animals (cows) older than two years. On an affected herd or flock basis, the animals may be grouped into four categories: clinical cases; asymptomatic silent shedders; subclinical carriers (non-shedders); and uninfected animals. Despite the fact that only 1 to 2 per cent of animals in a herd or flock may manifest clinical signs, up to 50 per cent or more of them may be infected (‘tip-of-iceberg’ or ‘pyramid’ phenomenon).27, 31, 121
A typical clinical case of bovine paratuberculosis is characterized by progressive weight loss and by continuous or intermittent chronic diarrhoea, which does not respond to treatment. (Figure 1) Alertness and appetite generally remain good.121 The faeces are soft to watery, are passed without straining, occasionally may contain casts of mucus, and are foul-smelling. The diarrhoea may decrease in severity during late pregnancy only to reappear more severely after parturition. The onset of the disease, particularly in high-producing animals, may be sudden. Milk production decreases, and the hair coat may become rough or areas of alopecia may develop. Submandibular oedema (bottle jaw) develops in some animals. Fever is usually absent. Terminally, there is severe dehydration, emaciation and weakness, and after a course of several weeks to months the animal dies. Although up to 50 per cent of the animals in a herd may be infected the annual mortality rate is usually less than 1 per cent. Secondary clinical complications associated with paratuberculosis can include increased susceptibility to mastitis and infertility.
In sheep and goats the disease is afebrile and is characterized by progressive emaciation. The consistency of the faeces may be normal throughout most of the course of disease, or it may be intermittently soft and pasty, and the pellets unformed. Terminally, the faeces are usually soft and pasty. Shedding of wool may occur in some sheep. The appetite is maintained throughout most of the clinical course of the disease. Subcutaneous oedema of the submandibular region may occur, but is rare.29

Pathology
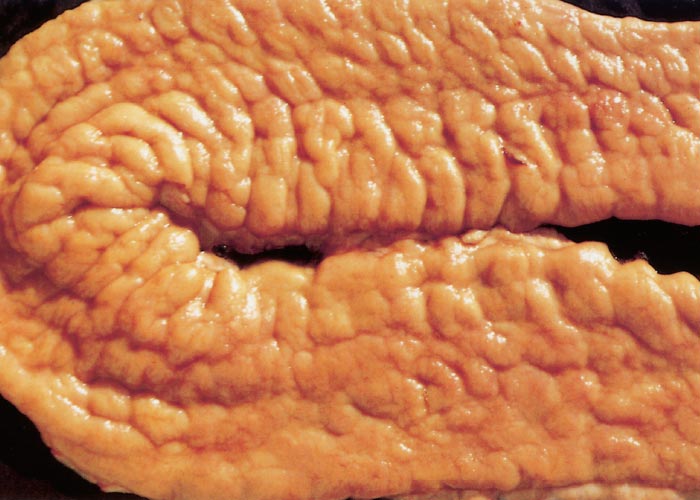
In clinical cases gross intestinal lesions compatible with those of paratuberculosis occur in more than 50 per cent of cattle22, 30, 121 and in 70 per cent of clinical cases in sheep.13 Gross changes in cattle and sheep are usually recognized in the mucosa of the distal jejunum and ileum, but in advanced ovine cases lesions may occur from the duodenum to the caecum.25 Diffuse thickening and corrugation of the intestinal mucosa is a characteristic finding in cattle. The mucosa forms stiff folds that do not disappear if the intestine is stretched and give it a convoluted appearance resembling the folds of the cerebrum (Figure 2). The intestinal serosal and mesenteric lymphatics are frequently inflamed, appearing prominent, beaded and cord-like (Figure 3).
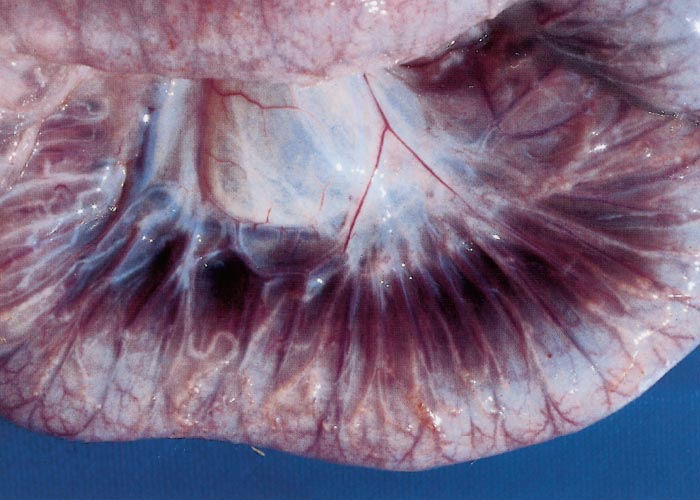
The ileocaecal and mesenteric lymph nodes are usually oedematous and severely enlarged.
As the disease progresses, oedema of the abomasal wall and serous fluid accumulations in the abdominal and pericardial cavities typify hypoproteinaemia. Pallor of the internal organs and mucous membranes reflects anaemia. Extra-intestinal organs, such as the liver, kidneys and lungs, may be involved on occasion.121 Only the kidneys may show macroscopic changes in these instances. In up to 25 per cent of clinically affected animals, calcification of the intima of the ascending aorta and the endocardium of the left atrium is present. Investigation into the pathogenesis of such mineralization is lacking in paratuberculosis, but comparative studies of non-caseating granulomatous diseases such as human sarcoidosis incriminated activated macrophages in the synthesis of vitamin D3 metabolites as side products in the deposition of calcium within the cardiovascular system.3
In infected goats, caseation with calcification has been observed in mesenteric lymph nodes. A recent publication described a granulomatous inflammation in medium-sized arteries in the intestinal submucosa and in the paracortex of mesenteric lymph nodes in animals experimentally infected with MAP.58 In some of these experimental animals ulceration of the intestinal mucosa over Peyers patches occurred. Intestinal strictures and fibrous adhesions have been noticed in some naturally infected goats.
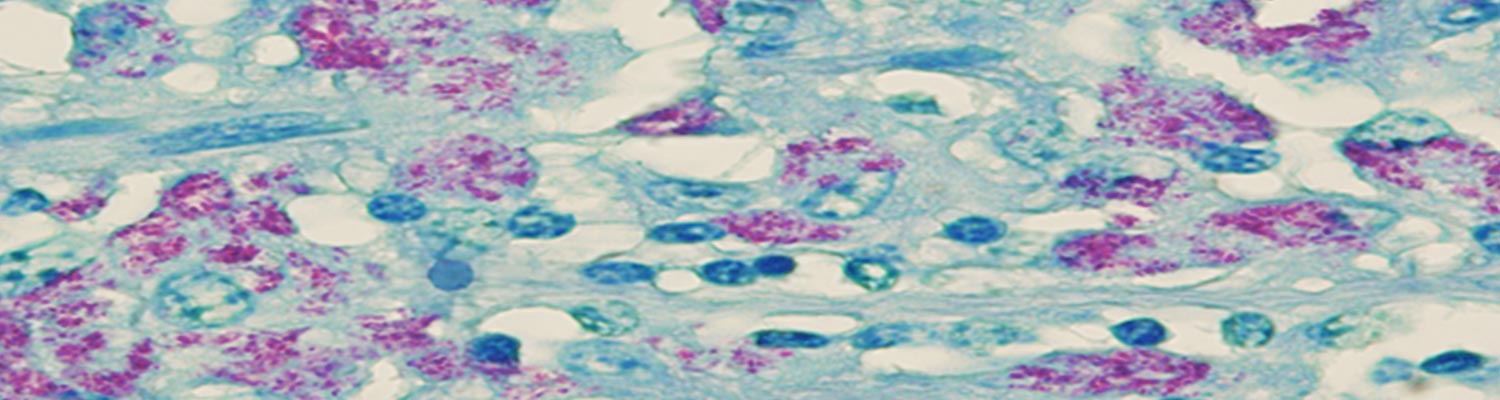
Microscopically, the characteristic lesion in clinical cases of paratuberculosis is non-caseating granulomatous inflammation, composed predominantly of epithelioid macrophages within the lamina propria and submucosa of the intestine. Secondary dilation of villous lacteals from inflammatory obstruction leads to protein-losing enteropathy. An infiltration of epithelioid macrophages completely replaces the paracortical zone of draining mesenteric lymph nodes.22 Single, isolated or numerous Langerhans type giant cells are intermingled with the epithelioid cells. Application of an acid-fast stain to sections of affected tissues usually enables the visualization of a plethora of engulfed bacilli within epithelioid and giant cells (Figure 4a), which is referred to as multibacillary (lepromatous) type of paratuberculosis and occurs most frequently in clinical cases. Rarely, the paucibacillary (tuberculous) form of paratuberculosis is encountered in animals with clinical disease. Scrapings or impression smears made from the mucosa of the intestine or from the cut surfaces of mesenteric lymph nodes can be stained with an acid-fast stain, such as Ziehl-Neelsen (ZN), to demonstrate clusters of the organisms.
Indirect immunohistochemistry can be applied to demonstrate the presence of MAP in the cytoplasm of infected epithelioid macrophages and Langerhans type giant cells. (Figure 4b). Whether this procedure is more sensitive than acid-fast staining is controversial.
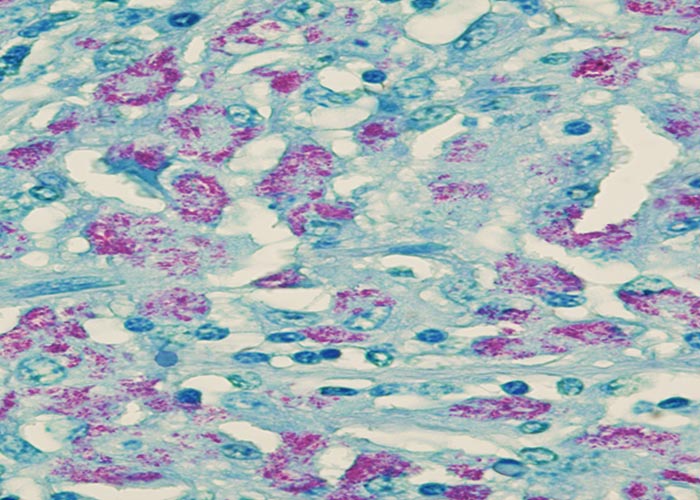
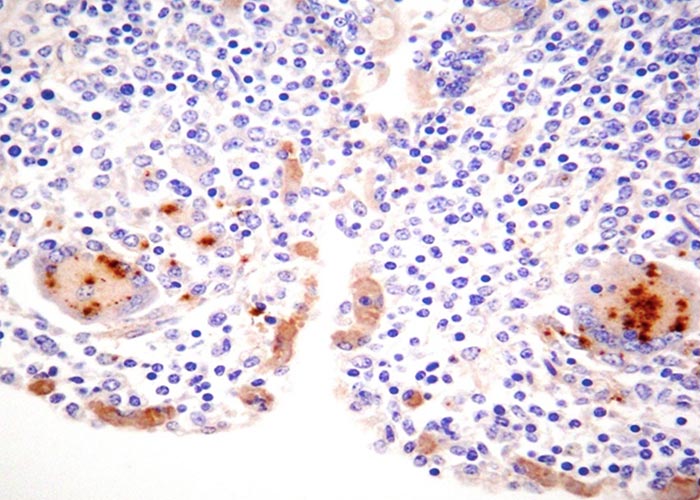
Microscopic involvement of the large intestine occurs to a much lesser degree; in both cattle and sheep the inflammatory response seldom extends beyond the ileocaecal valve. This observation is of significance when attempting to diagnose paratuberculosis with the help of rectal scrapings or rectal tissue taken for biopsy. The liver is a better organ of choice for biopsy than the rectum as it may contain small microgranulomas, which may occasionally contain acid-fast bacilli,22 but the technique is impractical to perform on a large scale.22 Similar microgranulomas may also occur in the lungs and kidneys.
Early lesions of paratuberculosis in subclinically affected cattle are characterized by the presence of individual Langerhans giant cells or a few scattered epithelioid macrophages in the tips of the intestinal villi or within the paracortical zone of the regional mesenteric lymph nodes and acid-fast bacilli are rarely detected.22
Ultrastructural studies revealed that the bacteria are located either within phagosomes or free within the perinuclear cytoplasm.13, 19
Diagnosis
A battery of diagnostic tests is available to identify the organisms directly (agent detection) or indirectly using specific immunological (serological) responses of the host.27, 32, 111 All tests currently employed suffer from a lack of sensitivity and specificity in that they fail to detect all infected animals and cannot discriminate between other mycobacterial or related bacterial infections that cross-react antigenically with MAP. A number of diagnostic tests are available that provide valuable supportive evidence for the presence of paratuberculosis. Because of the spectral aspect of the immune response to MAP in the host18 (i.e. fluctuating between cell-mediated immunity in the beginning of the infection to terminal humoral immunity) it is recommended that two or more tests be used simultaneously to confirm the diagnosis of the disease. The combination of tests should include a serological test, such as the AGID or enzyme-linked immunosorbent assay (ELISA), and/or the culture of faecal or tissue samples using liquid medium systems (see below) in conjunction with the PCR.
The available diagnostic tests and their combined characteristics as far as sensitivity (true infection) and specificity (true non-infection) are concerned are described below (see also Table 1).
For the purpose of culture from live animals a faecal sample of at least 10 g of faeces kept at 4 °C should reach the laboratory within four days of collection.86 If it is not possible to maintain specimens in transit at this temperature they should be submitted in a frozen state (−70 °C), but this reduces the viability of the organism.85 Mycobacterium avium subsp. paratuberculosis is extremely difficult to culture from the faeces of sheep even when bacteria can be demonstrated microscopically in acid-fast stained faecal smears.94
Table 1 Comparison of the results of diagnostic tests for paratuberculosis
| TEST | COMBINED SENSITIVITY (%) | SPECIFICITY (%) | TIME TO RUN |
| ELISA | 64 | 96 | 3 hrs |
| γ-INF | 70 | 97 | 28 hrs |
| E + γ | 92 | 96 | 28 hrs |
| CF | 26 | 96 | 3 hrs |
| AGID | 26 | 100 | 48 hrs |
| HEYM | 45 | 100 | 16 wks |
| BACTEC | 54 | 100 | 7 wks discontinued |
| M7H9C | Uk | 100 | 7 wks |
| PCR | 33 | 99 | 48 hrs |
| LN Biopsy | Uk | Uk | 48 hrs |
ELISA = Enzyme-linked immunosorbent assay
BACTEC = Radiometric culture system
γ-INF = Gamma interferon
PCR = Polymerase chain reaction
E + γ = ELISA plus gamma interferon
LN = Lymph node
CF = Complement fixation
AGID = Agar gel immunodiffusion plus ELISA HEYM = Herrold’s egg yolk medium
Uk = Unknown
Culture of MAP from faecal or tissue samples is the traditional diagnostic tool and is considered the ‘gold’ standard against which all other tests are compared.6 Numerous techniques (solid and liquid) for the primary cultivation of MAP from such samples have been developed. The original technique was to use Herrold’s egg-yolk medium (HEYM), a solid medium containing mycobactin, and to culture the organism at 37 °C for at least 12 weeks.6, 72 For the isolation of the organism from faeces or tissue specimens a decontamination procedure to destroy the spectrum of bacteria and fungi present other than the pathogenic mycobacteria must first be undertaken. The decontamination procedure which has given the best results is the treatment of the specimens with either benzalkonium chloride67 or hexadecylpyridinium chloride (HPC).119 Over the years, modifications of the techniques employed for specimen preparation have been developed.101
Other commercially available solid and liquid available media are Middlebrook 7H10, 7H11 and Middlebrook 7H9, respectively. In general, time for positive culture results is shorter when using liquid media.
A more rapid method for the detection of MAP was achieved by the use of a radiometric culture system in which a 14C-labelled substrate in a liquid medium containing egg-yolk is metabolized to C-labelled carbon dioxide in the gas phase above the culture.38 This method, known as Bactec 12B, has been discontinued since 2012 and is now replaced by an non-radioactive liquid medium known as M7H9C with similar high analytical sensitivity and cultivation potential for C and S strains of MAP.123 Confirmative readout of this technique must be combined with PCR as secondary test for direct confirmation of MAP. Incubation period for growth can be expected as early as 6 weeks, but the standard incubation period of 12 weeks is recommended to avoid false-negative results.
A faster alternative approach to direct agent detection is the gene amplification technique based on DNA analysis.112 The diagnostic specificity for this method is considered to be 97 to 100 per cent. The disadvantages of this test are its low sensitivity (35 per cent) and high cost and requirement for skilled technical help and certain equipment. The development of the PCR, which is based on DNA homology, has enabled rapid identification of MAPwithin three days.5, 116 Probes containing repetitive DNA elements are used to measure genetic relatedness among isolates of the same species. The detection of the IS900, a repetitive DNA insertion element of 1,5 kb present in 15 to 20 copies of theMAP genome, was a milestone in the search for a genetic marker for this pathogen.15, 65 The IS900 genome has a molecular size of 4,4–4,7 × 106 bp25 and a unique nucleotide sequence that can be specifically detected by PCR or hybridization.52, 65 IS900 and IS902 have been used for the taxonomic studies of MAP and M. avium isolates, respectively. Sequences related to IS900 are also found in wood pigeon mycobacteria (IS902) and M. avium (IS901).112, 114 A comparison of 16S rRNA sequences has shown that the 16S rRNA of MAP is 99,9 per cent homologous to M. avium and 98,1 per cent homologous to Mycobacterium bovis.114 Furthermore, the use of DNA fingerprinting techniques based on IS900 as a target sequence test has been particularly successful in characterizing MAP strains.33 Application of the restriction fragment length polymorphism (RFLP) technique, which uses restriction endonuclease and DNA hybridization analyses of various MAP isolates, has revealed a considerable degree of genetic variability between several MAP isolates, and makes the identification of individual strains of clonal lineage possible.41, 75, 76, 120 These techniques are useful in the retrospective tracing of the origin of strains in cases of dispute concerning the introduction of MAP into a herd previously free of the infection. Identification of MAP strains allows not only the monitoring of both horizontal and vertical spread of infection, but also an analysis of the origin of imported infected animals and of the transmission of infection among farm and wild ruminants.88 An additional species-specific recombinant clone, F57, has been generated in a transcription vector and sequenced.28, 82 Its sequence has revealed that it is unique and unlike any other known gene including IS900. Another probe that is based on 16S rRNA has been developed for MAP identification.115, 120 Since rRNA is present in the organisms, rRNA-derived detection methods for MAP are likely to prove useful in identifying MAP -infected animals. The 5S rRNA gene has been selected to examine species and subspecies differentiation of related mycobacteria including MAP and M. avium.27
Histopathological examination of mesenteric lymph node or intestinal biopsy specimens is a further method for direct visualization of the pathogen.20, 37, 78 Characteristic lesions of paratuberculosis are most prominent in the paracortex of draining mesenteric lymph nodes.78 Tissue sections are stained with the haematoxylin and eosin and ZN stains (Figure 4a) or, alternatively, are processed immunohistochemically (Figure 4b) prior to microscopic examination. A less laborious alternative to intestinal or mesenteric lymph node biopsies is the examination of percutaneously obtained liver biopsies.22 The examination of rectal biopsy specimens or smears made from scrapings of the rectal mucosa is extremely unreliable, as specific lesions or organisms are rarely found in the rectal wall.
The benefits of the immunohistochemical demonstration of MAP antigen in macrophages and giant cells are known.32, 81, 102 The value of this test is that it enables free antigens, cell fragments and fragmented cell walls to be detected, in contrast to the histopathological examination of tissues stained by an acid-fast method in which only organisms that were viable prior to the collection of the specimens can be detected. Tissues obtained by biopsy can also be subjected to DNA amplification methods such as PCR.70, 81
The criteria currently accepted for a positive histological diagnosis of Johne’s disease are the presence of epithelioid and/or Langerhans-type giant cells in the tissues and the demonstration of acid-fast bacilli in these cells. The latter is not always possible in animals in the very early stages of Johne’s disease in which only a few acid-fast bacilli may be present, or in cases of the paucibacillary form of paratuberculosis. In these cases the immunohistochemical procedures may be more effective.
Direct microscopic examination of faecal smears stained by the ZN method is an inexpensive, rapid method to identify infected animals that excrete large numbers of bacilli. The bacilli typically occur in clumps of 4 to 20 or more. In order to detect them directly in such smears the faeces must contain a minimum number of 104 colony-forming units per gram. The presence of large saprophyte acid-fast bacilli may complicate the interpretation of the results of such examinations.
The three commonly used serological tests to detect antibodies directed against MAP are the complement fixation (CF), AGID, and absorbed ELISA.88, 99 Results of serological tests on individual animals are often inconclusive, but they are of value when performed on a herd/flock basis as an indicator of exposure to the organism. False positive reactions in the serological tests may be due to cross reactions with other micro-organisms, as the antigens used in the tests are crude and contain antigens common to the majority of bacteria of the order Actinomycetales. False-negative responses may occur due to an insufficient antibody level (too low a titre in early infection) or when a state of anergy develops in clinical cases attributable to the protein-losing enteropathy, which includes the loss of immunoglobulins from severely affected intestines.
The CF test is used to certify animals free of infection for export/import purposes. However, it has low sensitivity and specificity as it is not very species-specific, is dependent on a cut-off point, and is cross-reactive with bacteria belonging to the genera Trueperella, Nocardia and Rhodococcus.45, 66, 124 The AGID is a serological test with a high antibody detection limit (e.g. clinical cases).97, 98 It is a simple assay and results are available within 48 hours. The test is inexpensive to conduct and is 100 per cent specific, but has a low sensitivity of between 17 and 40 per cent for subclinically infected animals and 56 per cent for clinical cases.68
Low antibody titres can be detected by application of the absorbed ELISA test and therefore some cattle in the subclinical stage of the disease can be identified. Its sensitivity can be improved by pre-absorption with Mycobacterium phlei.128 The sensitivity of this test may be as low as 15 per cent in low-level faecal shedders whereas in animals with clinical disease sensitivity can be as high as 87 per cent. Results depend on the determination of the cut-off point89 and the diagnostic test used as a standard.108 Combined analyses of data indicate that the sensitivity for the absorbed ELISA is about 45 per cent when applied to cattle herds consisting only of animals in the preclinical stage of infection. The specificity of the ELISA is 99,7 per cent. It is relatively inexpensive to perform and commercially produced ELISA kits are available.34 The principal factor determining the efficacy of the ELISA depends on the type of antigen used, a lipoarabinomannan (LAM),105 a recombinant polypeptide (a362),31 a purified specific antigen,1 or a crude whole cell extract (which is used in most laboratories). The ELISA can also be conducted on milk samples. There is no correlation, however, between antibody titres in the milk and those in the serum of the same animal.48
A novel ELISA has been introduced as ethanol-vortex enzyme-linked immunosorbent assay (EVELISA) with MAP surface antigens dislodged by treatment with 80 per cent ethanol followed by a 30 second agitation by vortex.39 The diagnostic specificity and sensitivity are listed as 97.4 and 100 per cent, respectively.
Three cell-mediated immunity tests are available: the skin sensitivity test, the lymphocyte transformation test and the gamma interferon test. These are based on the premise that infected animals first develop cell-mediated immunity from epitopes produced by MAP which, upon challenge of the animal, can be measured. The skin sensitivity test which, although used for many years for the diagnosis of paratuberculosis, is rarely performed these days.27 It should be conducted during the preclinical stage of the disease as it is of little value once clinical signs appear. A positive skin reaction does not provide sufficient evidence to confirm the presence of infection with the causative organism of paratuberculosis, as cross-reactivity with other pathogenic and saprophytic Mycobacteria spp. occurs, and a positive reaction may also be obtained in cattle previously vaccinated against paratuberculosis. The lymphocyte transformation test is based on the in vitro detection of specifically sensitized lymphocytes.27, 40 It is labour intensive, requires a reasonably sophisticated laboratory, depends on the use of radioactive substances and must be performed on fresh blood samples. A variation and spin-off of the lymphocyte transformation test is the gamma interferon test.21, 127 It measures γ-interferon, which is a cytokine produced by specifically sensitized lymphocytes.127 Mycobacterium avium is used in the test as a stimulogen. It is technically not easy to perform and must be performed on living and preserved cells, but has the potential to identify subclinical cases in the early stages of infection. The sensitivity of the test is in the region of 70 per cent and the specificity approximately 97 per cent. Although it is commercially available as a kit there are few data in the literature on the efficiency of the gamma interferon test in the diagnosis of paratuberculosis.127
When choosing one or a combination of tests it is important to recognize the spectral aspect of the immune response in paratuberculosis and the costs involved. Cost factors limit the use of a number of tests as herd tests.35 In order to establish a definitive diagnosis of paratuberculosis in a herd, agent detection methods should be chosen as well as a confirmatory necropsy. By virtue of their low costs, serological tests are suitable as herd tests. A sensitive, practical and economical way of diagnosing paratuberculosis is to perform a combination of an ELISA and strategically pooled faecal culture followed by PCR.24, 122
Differential diagnosis
Emaciation, profuse diarrhoea and weakness are seen in typical cases of paratuberculosis in cattle manifesting clinical signs and differential diagnoses hence include conditions causing similar clinical signs such as gastrointestinal endoparasitism, in particular paramphistomosis and schistosomosis, bovine virus diarrhoea/mucosal disease, enzootic bovine leukosis, salmonellosis, coccidiosis and renal amyloidosis. In sheep differential diagnoses include conditions leading to wasting and emaciation such as rumenitis, coccidiosis, chronic verminosis and inadequate feeding during winter or in times of drought, as well as marasmus due to cobalt deficiency.
Control
Chemotherapy with tuberculostatic drugs is both impractical and ineffective in animals suffering from paratuberculosis and as a result control measures consist primarily of herd management/husbandry and vaccination strategies. Numerous national and state paratuberculosis control programmes have been employed throughout the world that utilize a combination of management practices and/or vaccination.17, 87
The practice of controlling paratuberculosis through testing and culling has been widely recommended and is used in many parts of the world.34 Diagnostic tests for paratuberculosis are improving, but accurate detection of all infected animals, especially those in the early course of infection and transmitting the organism within a herd, is still not possible. This fact makes test and cull strategies somewhat ineffective in eradication, except when only targeting high-shedding animals in the herd.14, 63 Therefore, it is widely recognized that unless animals can be detected early during infection, vaccination likely remains the best hope for controlling and preventing the disease.9 Control through management practices on a herd or flock basis depends on the elimination of infected animals and strict attention to hygiene and other husbandry procedures to prevent spread of infection. Many of the prophylactic measures employed are aimed at neonatal calves, as young animals are the most susceptible to the infection. It is recommended that the milk of infected cows should not be fed to calves, as the causative organism can be isolated from the milk of clinically and subclinically infected cattle.109 A paratuberculosis-free status should be the ultimate goal, with maintenance of that status. Water, feed and pasture should be regularly assessed for the presence of MAP by means of PCR or culture methods.17, 87
Although commercial vaccines are available, control of paratuberculosis by vaccination alone is currently not an effective eradication method.56, 79 Vaccination is regulated in some countries/states and should only be performed in tuberculosis-free herds, as inoculation against paratuberculosis in areas where tuberculosis jointly occurs may interfere with results obtained in the tuberculin skin test for the diagnosis of tuberculosis. Currently, there are at least three commercial paratuberculosis vaccines worldwide, but only one (Mycopar®) is approved for limited use in the US.14 Silirum® is produced by Zoetis. Mycopar® is manufactured by Boehringer Ingelheim Vetmedica Inc. whereas Gudair® is produced by CZ Veterinaria in Spain and is being used in countries with large sheep populations.14 Importantly, all three consist of an inactivated whole-cell preparation of either a strain of M. avium subspecies avium, also known as Paratuberculosis Strain 18 (Mycopar®), or M. avium subspecies paratuberculosis strain 316F (Silirum® and Gudair®).9 The ideal vaccine would completely prevent infection and/or promote protective immunity, thus blocking both horizontal and vertical transmission. Current vaccines for paratuberculosis fall far short of this high standard. The vaccines have been shown to be effective at lowering faecal shedding levels,42, 50, 55 tissue colonization50, 107 or clinical disease incidence,50, 104 but do not completely eliminate all three. The use of purified proteins, a DNA vaccine expressing proteins or an oral attenuated mutant vaccine able to induce both a cellular immune response and protection from disease would be a realistic prophylactic approach with less potential effect on tuberculosis testing and reduced administration site reactions. Ideally, different proteins should be used for vaccination and for diagnostic tests.40 Currently, there are no subunit, live mutant or DNA vaccines commercially available against paratuberculosis.9 A few subunit vaccines, DNA vaccines and several live knockout mutant vaccines have been tested experimentally in mice and/or goats that demonstrated some efficacy in reducing shedding, pathology and/or clinical disease, but did not prevent infection.9
Genetic factors may affect the susceptibility of animals to paratuberculosis, but as yet specific genes governing susceptibility or resistance to the disease have not been fully elucidated. A gene has been isolated from mice and designated Nramp1 (natural resistance-associated macrophage protein) gene.117 It has been suggested that Nramp1 may play a role in determining resistance to mycobacteria in mice. The Nramp1 gene has now been renamed SLC11A1 (Solute Carrier Family 11 Member 1).7 Recently, the caprine SLC11A1 gene has been shown to have a significant role in paratuberculosis resistance in goats.2 Mutations in the SLC11A1 gene in humans have been associated with susceptibility to infectious diseases such as tuberculosis and leprosy, and numerous inflammatory diseases including rheumatoid arthritis and Crohn's disease.7 In future it may be possible to select genetically resistant animals to aid in the control and/or eradication of paratuberculosis.
Paratuberculosis as a Potential Public Health Danger and its Relationship with Crohn’s Disease
Crohn’s disease is a chronic relapsing inflammatory disease of humans affecting any part of the intestinal tract, but in particular the terminal ileum or colon, or both. It is more common in Western countries, where its prevalence has been increasing over the last few decades. It usually affects adolescents or young adults, with a secondary peak in elderly people. The clinical signs vary depending on the site of inflammation, patients with predominantly ileal lesions having symptoms resembling acute appendicitis whilst those with predominantly colonic lesions develop abdominal pain and diarrhoea, often with haemorrhage. Pathologically, the disease is characterized by non-caseating transmural granulomatous enteritis with extensive ulceration and fistulation.96 Although acid-fast organisms have not been detected histologically in the gut of affected individuals, ultrastructural studies have revealed the presence of ZN-negative, cell wall-deficient organisms in the lesions of several patients with Crohn’s disease.28, 49 Mycobacterium avium subsp. paratuberculosis has been isolated from only a small proportion of patients by conventional cultural methods, but in a survey it was identified from the gut of 86 per cent of patients by the use of improved liquid culture methods and PCR techniques.49
As MAP is widely prevalent in the intestinal tract of various farmed livestock and wildlife species and can survive for extended periods in the environment, it has the potential to contaminate pastures and subsequently human water supplies.49, 59 The occurrence of clustered cases of Crohn’s disease in 11 Cardiff County towns in the UK is believed to have been caused by contamination of the water supply of these towns during the patients’ childhood years.49
Several studies have demonstrated the thermo-resistance of MAP to the traditional milk pasteurization process,26, 47, 51, 69, 100 and an estimated 35 per cent of clinically infected cattle and 12 per cent of subclinically infected cows have detectable numbers of MAP in their milk.28 As certain humans appear to have an inherited or acquired sensitivity to Crohn’s disease, it is speculated that cumulative exposure to MAP by these persons via prolonged ingestion of either contaminated water or milk can, as in animals, result in colonization of the ileum and colon.73 Subsequent exposure to a trigger factor(s), such as intercurrent infection or stress, may then result in the development of the disease.
Given these circumstances, control of paratuberculosis for food safety should be a desirable goal.118
References
- ABBAS, B., RIEMANN, H. P. & LOMMERDAL, B., 1983. Isolation of specific peptides from Mycobacterium paratuberculosis protoplasm and their use in an enzyme-linked immunosorbent assay for the detection of paratuberculosis in cattle. American Journal of Veterinary Research, 44, 2229–2236.
- ABRAHAM, A., NAICY, T., RAGHAVAN, K. C., SIJU, J. & ARAVINDAKSHAN, T., 2017. Evaluation of the association of SLC11A1 gene polymorphism with incidence of paratuberculosis in goats. Journal of Genetics, 96(4), 641-646.
- ADAMS, J. S. & GACAD, M. A., 1985. Characterization of 1-alpha Hydroxylation of Vitamin D Sterols by Cultured Alveolar Macrophages from Patients with Sarcoidosis. Journal Experimental Medicine, 161, 755-765.
- ALZUHERRI, H. M., LITTLE, D. & CLARKE, C. J., 1997. Altered intestinal macrophage phenotype in ovine paratuberculosis. Research in Veterinary Science, 63, 139–143.
- AMBROSIO, R. E., HARRIS, Y. & HUCHZERMEYER, H. F. A. K., 1991. A DNA probe for the detection of Mycobacterium paratuberculosis. Veterinary Microbiology, 26, 87–93.
- ANON., 1985. Laboratory methods in veterinary mycobacteria. The Mycobacteriology Unit, National Veterinary Services, Animals and Plant Health Inspection Service. US Department of Agriculture, Ames, Iowa, 55-57.
- AWOMOYI, A. A., 2007. The human solute carrier family 11 member 1 protein (SLC11A1): linking infections, autoimmunity and cancer. FEMS Immunology and Medical Microbiology, 49(3), 324–329.
- BANG, B., 1906. Chronische pseudotuberkuloese Darmentzuendung beim Rind. Berliner Tierärztliche Wochenschrift, 42, 759–763.
- BANNANTINE, J. P., HINES, M. E. I., BERMUDEZ, L. E., TALAAT, A. M., SREEVATSAN, S., STABEL, J. R., CHANG, Y. F., COUSSENS, P. M., BARLETTA, R. G., DAVIS, W. C., COLLINS, D. M., GRÖHN, Y. T. & KAPUR, V., 2014. A rational framework for evaluating the next generation of vaccines against Mycobacterium avium subspecies paratuberculosis. Frontiers in Cellular Infection Microbiology, 9(4), 126. doi: 3389/fcimb.2014.00126.
- BANNENTINE, J. P. & BERMUDEZ, L. E., 2013. No Holes Barred: Invasion of the Intestinal Mucosa by Mycobacterium avium subsp. paratuberculosis. Infection and Immunity, 81, 3960-3965.
- BARKEMA, H. W., HESSELINK, J. W., MCKENNA, S. L. B., BENEDICTUS, G. & GROENENDAAL, H., 2010. Global Prevalence and Economics of Infection with Mycobacterium avium subspecies paratuberculosis. In: BEHR, M & COLLINS, D., (eds.). Chapter 2. Paratuberculosis: Organism, Disease and Control. CABI, Oxfordshire, UK. ISBN: 978 1 84593 613 6.
- BASSEY, E. O. E. & COLLINS, M. T., 1997. Study of T-lymphocyte subsets of healthy and Mycobacterium avium subsp. paratuberculosis-infected cattle. Infection and Immunity, 4869–4872.
- BASTIANELLO, S. S., 1989–1998. Ruakura Animal Health Laboratory, Agriquality, East Street, Hamilton, New Zealand. Unpublished observations.
- BASTIDA, F. & JUSTE, R. A., 2011. Paratuberculosis control: a review with a focus on vaccination. Journal of Immune Based Therapeutic Vaccines, 9, 8. doi: 10.1186/1476-8518-9-8.
- BAUERFEIND, R., BENAZZI, S., WEISS, R., SCHLIESSER, T., WILLEMS, H. & BALJER, G., 1996. Molecular characterization of Mycobacterium paratuberculosis isolates from sheep, goats, and cattle by hybridization with a DNA probe to insertion element IS900. Journal of Clinical Microbiology, 34, 1617–1621.
- BEARD, P., RHIND, S., BUTON, D., DANIELS, M. J., HENDERSON, D., PIRIE, A., RUDGE, K., GRAIG, A., HUTCHINGS, M. A., STEVENSON, K. & SHARP, J. M., 2001. Natural paratuberculosis infection in rabbits in Scotland. Journal of Comparative Pathology, 124, 290-299.
- BEHR, M. A. & COLLINS, D. M., 2010. Paratuberculosis. Organism, Disease, Control. CABI, Oxfordshire, UK. ISBN: 978 1 84593 613 6.
- BENDIXEN, P. H., 1978. Immunologic reactions caused by infection with Mycobacterium paratuberculosis. A review. Nordisk Veterinaermedicin, 30, 163–168.
- BENDIXEN, P. H., BLOCH, B. & JORGENSEN, J. B., 1981. Lack of intracellular degradation of Mycobacterium paratuberculosis by bovine macrophages infected in vitro and in vivo: light microscopic and electron microscopic observations. Amercian Journal of Veterinary Research, 42, 109–113.
- BENEDICTUS, G. & HAAGSMA, J., 1986. The efficacy of mesenteric lymph node biopsy in the eradication of paratuberculosis from an infected dairy farm. Veterinary Quarterly, 8, 5-11.
- BILLMAN-JACOBE, H., CARRIGAN, M., COCKRAM, F., CORNER, L. A., GILL, I. J., HILL, J. F., JESSEP, T., MILNER, A. R. & WOOD, P. R., 1992. A comparison of the interferon gamma assay with the absorbed ELISA for the diagnosis of Johne’s disease in cattle. Australian Veterinary Journal. 69, 25-28.
- BUERGELT, C. D., HALL, C., MCENTEE, K. & DUNCAN, J. R., 1978. Pathological evaluation of paratuberculosis in naturally infected cattle. Veterinary Pathology, 196–207.
- BUERGELT, C. D., LAYTON, A. W., GINN, P. E., TAYLOR, M., KING, J. M., HABECKER, P. L., MAULDIN, E., WHITLOCK, R., ROSSITER, C. & COLLINS, M. T., 2000. The pathology of spontaneous paratuberculosis in the North American bison (Bison bison). Veterinary Pathology, 428–438.
- BULAGA, L. L. & COLLINS, M. T., 1999. U.S. Voluntary Johne’s Disease Herd Status Program for Cattle. USAHA National Johne’s Disease Working Group Certification Subcommittee. Proceedings of the Sixth International Colloquium on Paratuberculosis. 14–18 February 1999. Melbourne, Australia, 39–47.
- CARRIGAN, M. J. & SEAMAN, J. T., 1990. The pathology of Johne’s disease in sheep. Australian Veterinary Journal, 67, 47-50.
- CHIODINI, R. J. & HERMAN-TAYLOR, J., 1993. The thermal resistance of Mycobacterium paratuberculosis in raw milk under conditions simulating pasteurization. Journal of Veterinary Diagnostic Investigation, 5, 629–631.
- CHIODINI, R. J., VAN KRUININGEN, H. J. & MERKAL, R. S., 1984. Ruminant paratuberculosis (Johne’s disease): The current status and future prospects. Cornell Veterinarian, 74, 218–262.
- CHIODINI, R. J., VAN KRUININGEN, H. J., THAYER, W. R., MERKAL, R. S. & COUTU, J. A., 1984. Possible role of mycobacteria in inflammatory bowel disease. Digestive Diseases and Sciences, 29, 1073–1079.
- CLARK, C. J., 1997. The Pathology and Pathogenesis of Paratuberculosis in ruminants and other species. Journal Comparative Pathology, 116, 217-226.
- CLARKE, C. J., 1997. The pathology and pathogenesis of paratuberculosis in ruminants and other species. Journal of Comparative Pathology, 16, 217–261.
- COCITO, C., GILOT, P., COENE, M., DEKESEL, M., POUPART, P. & VANNUFFEL, P., 1994. Paratuberculosis. Clinical Microbiology Review, 7, 328–345.
- COETSIER, C., HAVAUX, X., MATTELARD, F., SADATTE, S., CORMONT, F., BUERGELT, C., LIMBOURG, B., LATINNE, D., BAZIN, H., DENEF, J. F. & COCITO, C., 1998. Detection of Mycobacterium avium subsp. paratuberculosis in infected tissues by new species-specific immunohistological procedures. Clinical and Diagnostic Laboratory Immunology, 5, 446– 451.
- COLLINS, D. M., CAVAIGNAC, S. & DELISLE, G., 1997. Use of four DNA insertion sequences to characterize strains of the Mycobacterium avium complex isolated from animals. Molecular and Cellular Probes, 11, 373–380.
- COLLINS, M. T., ANGULO, A., BUERGELT, C. D., HENNAGER, S. G., HIETALA, S. K., JACOBSON, R. H., WHIPPLE, D. L. & WHITLOCK, R. H., 1993. Reproducibility of a commercial enzyme-linked immunosorbent assay for bovine paratuberculosis among eight laboratories. Journal of Veterinary Diagnostic Investigation, 5, 52-55.
- COLLINS, M. T. & MORGAN, I. R., 1991. Economic decision analysis model of a paratuberculosis test and cull program. Journal of the American Veterinary Association, 199, 1724–1729.
- COLLINS, M. T., TEUBERT, D. G. & OSTERHUIS, J. E., 1994. Paratuberculosis in 28 species at the San Diego Wild Animal Park ( abstract). Proceedings of the Fourth International Colloquium on Paratuberculosis. 17–21 July 1994. Cambridge, UK, 32-34.
- CONDRON, R., SCHROEN, C., BLACK, C., RIDGE, S. E. & HOPE, A., 1994. Histological confirmation of subclinical infection with Mycobacterium paratuberculosis in cattle.Proceedings of the Fourth International Colloquium on Paratuberculosis.17–21 July 1994. Cambridge, UK, 37-40.
- COUSINS, D. V., EVANS, R. J. & FRANCIS, B. R., 1995. Use of BACTEC radiometric culture medium and polymerase chain reaction for the rapid screening of ruminant faeces and intestinal tissues for Mycobacterium paratuberculoisis. Australian Veterinary Journal, 72, 458-467.
- EDA, S., BANNANTINE, J. P., WATERS, W. R., MORI, Y., WHITLOCK, R. H., SCOTT, M. C. & SPEER, C. A., 2006. A highly sensitive and subspecies-specific surface antigen enzyme-immunosorbent assay for diagnosis of Johne’s disease. Clinical Vaccine Immunology, 13, 837-844.
- EMERY, D. L., DUFFY, J. H. & WOOD, P. R., 1988. An analysis of cellular proliferation and synthesis of lymphokines and specific antibody in vitro by leukocytes from immunized cattle. Veterinary Immunology Immunopathology, 18, 67–80.
- ERIKS, I., MUNCK, K., BESSER, T. E., CANTOR, G. H. & KAPUR, V., 1996. Rapid differentiation of Mycobacterium avium and M. paratuberculosis by PCR and restriction enzyme analysis. Journal of Clinical Microbiology, 34, 734–737.
- FAISAL, S. M., CHEN, J. W., YAN, F., CHEN, T. T., USEH, N. M., GUO, S., YAN, W., WANG, S. J., GLASER, A. L., MCDONOUGH, S. P., SINGH, B. & CHANG, Y. F., 2013. Evaluation of a Mycobacterium avium subsp. paratuberculosis leuD mutant as a vaccine candidate against challenge in a caprine model. Clinical Vaccine Immunology, 20, 572–581. doi: 10.1128/CVI.00653-12.
- FECHNER, K., MAETZ-RENSING, K., LAMPE, K., KAUP, F., CZERNY, C. P. & SCHAEFER, J., 2017. Detection of Mycobacterium avium subsp. paratuberculosis in non-human primates. Journal Medical Primatology, 46, 211-217.
- FECTEAU, M. E., 2018. Paratuberculosis in Cattle. Veterinary Clinics of North America Food Animal Practice, 34(1), 209-222.
- GILMOUR, N. J. L. & GOUDSWAARD, J., 1972. Corynebacterium renale as a cause of reactions to the complement fixation test for Johne’s disease. Journal of Comparative Pathology, 82, 333–336.
- GILMOUR, N. J. L. & NYANGE, J., 1989. Paratuberculosis (Johne’s disease) in deer. Practice, 11, 193–196.
- GRANT, I. R., BALL, H. J., NEILL, S. D. & ROWE, M. T., 1996. Inactivation of Mycobacterium paratuberculosis in cow’s milk at pasteurization temperatures. Applied and Environmental Microbiology, 62, 631–636.
- HARDIN, L. E. & THORNE, J. G., 1996. Comparison of milk with serum ELISA for the detection of paratuberculosis in dairy cows. Journal of the American Veterinary Association, 209, 120–122.
- HERMON-TAYLOR, J., 2000. Mycobacterium avium subspecies paratuberculosis in animals, food products and in water supplies and its impact on Human Health. Cattle Practice: Journal of the British Cattle Veterinary Association, 8, 355–360.
- HINES, M. E., II., TURNQUIST, S. E., ILHA, M. R. S., RAJEEV, S., JONES, A. L., WHITTINGTON, L., BANNANTINE, J. P., BARLETTA, R. G., GRÖHN, Y. T., KATANI, R., TALAAT, A. M., LI, L. & KAPUR, V., 2014. Evaluation of Novel Oral Vaccine CR Gandidates and Validation of a Caprine Model of Johne’s Disease. Frontiers in Cellular and Infection Microbiology, 4, 1014. doi: 10.3389/fcimb.2014.00026.
- HOPE, A. F., TULK, P. A. & CONDRON, R. J., 1996. Pasteurisation of Mycobacterium paratuberculosis in whole milk. Proceedings of the Fifth International Colloquium on Paratuberculosis. 29 September – 4 October 1996. Madison, Wisconsin, 377-382.
- HURLEY, S. S., SPLITTER, G. A. & WELCH, R. A., 1989. Development of a diagnostic test for Johne’s disease using DNA hybridization probe. Journal of Clinical Microbiology, 27, 1582–1587.
- JOHNE, H. A. & FROTHINGHAM, L., 1895. Ein eigenthuemlicher Fall von Tuberkulosis beim Rind. Deutsche Zeitschrift Für Tiermedizinische und Vergleichende Pathologie, 21, 438–454.
- JOHNSON-IFEARULUNDI, Y. J. & KANEENE, J. B., 1997. Relationship between soil type and Mycobacterium paratuberculosis. Journal of the American Veterinary Medical Association, 210, 1735–1740.
- KALIS, C. H., HESSELINK, J. W., BARKEMA, H. W. & COLLINS, M. T., 2001. Use of long-term vaccination with a killed vaccine to prevent fecal shedding of Mycobacterium avium subsp paratuberculosis in dairy herds. American Journal of Veterinary Research, 62, 270-274. doi: 10.2460/ajvr.2001.62.270.
- KOERMENDY, B., 1994. The effect of vaccination on the prevalence of paratuberculosis in large dairy herds. Veterinary Microbiology, 41, 117-125.
- KOPECKY, K. E., 1977. Distribution of paratuberculosis in Wisconsin, by soil regions. Journal of the American Veterinary Association, 170, 320–324.
- KRUEGER, C., KOEHLER, H. & LIEBLER-TENORIO, E. M., 2015. Sequential development of lesions 3,6,9 and 12 months after experimental infection of goat kids with Mycobacterium avium subspecies paratuberculosis. Veterinary Pathology, 52, 276-290.
- LARSEN, A. B., MERKAL, R. S. & VARDAMAN, T. H., 1956. Survival time of Mycobacterium paratuberculosis. American Journal of Veterinary Research, 39, 65-69.
- LARSEN, A. B., STALHEIM, O. H. V., HIGHES, D. E., APPEL, L. H., RICHARDS, W. D. & HIMES, E. M., 1981. Mycobacterium paratuberculosis in the semen and genital organs of a semen-donor bull. Journal of the American Veterinary Association, 179, 169-171.
- LEPPER, A. W. D. & WILKS, C. R., 1988. Intracellular iron storage and the pathogenesis of paratuberculosis. Comparative studies with other mycobacterial, parasitic or infectious conditions of veterinary importance. Journal of Comparative Pathology, 98, 31-53.
- LITTLE, D., CLARK, C. J. & ALZUHERRI, H. M., 1994. Phenotypic characterization of intestinal lymphocytes in ovine paratuberculosis. Proceedings of the Fourth International Colloquium on Paratuberculosis. 17–21 July 1994, Cambridge, UK, 168–171.
- LU, Z., BRANSCUM, A. J., SHUCK, K. M., ZHANG, J., DUBOVI, E. J., TIMONEY, P. J. & BALASURIYA, U. B. R., 2008. Comparison of two real-time reverse transcription polymerase chain reaction assays for the detection of Equine arteritis virus nucleic acid in equine semen and tissue culture fluid. Journal of Veterinary Diagnostic Investigation, 20(2), 147-155.
- MANNING, E. J. B., STEINBERG, H., ROSSOW, K., RUTH, G. R. & COLLINS, M. T., 1998. Epizootic of paratuberculosis in farmed elk. Journal of the American Veterinary Association, 213, 1320–1322.
- MCFADDEN, J. J., GREEN, E. & HERMON-TAYLOR, J., 1988. DNA probes to identify and detect Mycobacterium paratuberculosis in clinical and veterinary samples. Proceedings of the Second International Colloquium on Paratuberculosis. 22 and 23 September 1988. Maisons-Alfort, France, 201–205.
- MCKENZIE, R. A. & WARD, W. H., 1981. Rhodococcus (Corynebacterium) equi: A possible cause of reactions to the complement fixation test for Johne’s disease in cattle. Australian Veterinary Journal, 57, 200-201.
- MERKAL, R. S. & THURSTON, J. R., 1968. Susceptibilities of mycobacterial and nocardial species to benzalkonium chloride. American Journal of Veterinary Research, 29, 759–761.
- MICHEL, A., 1999. Ovine paratuberculosis in South Africa—prevalence and serological diagnosis. Abstracts of the Laboratory Diagnosticians Group of the South African Veterinary Association/State Veterinarians. August 1997. Journal of the South African Veterinary Association, 70, 54-55.
- MILLAR, D., FORD, J., SANDERSON, J., WHITHEY, S., TIZARD, M., DORAN, T. & HERMON-TAYLOR , J., 1996. IS900 PCR to detect Mycobacterium paratuberculosis in retail supplies of whole pasteurized cow’s milk in England and Wales. Applied and Environmental Microbiology, 62, 3446–3452.
- MILLER, J., JENNY, A., RHYAN, J., SAARI, D. & SUAREZ, D., 1997. Detection of Mycobacterium bovis in formalin-fixed, paraffin-embedded tissues of cattle by PCR amplification of an IS6110 sequence specific for Mycobacterium bovis complex organisms. Journal of Diagnostic Investigation, 9, 244-249.
- MOMOTANI, E., WHIPPLE, D. L., THIERMANN, A. B. & CHEVILLE, N. F., 1988. Role of M-cells and macrophages in the entrance of Mycobacterium paratuberculosis into domes of ileal Peyer’s patches of calves. Veterinary Pathology, 25, 131–137.
- MORRISON, N. E., 1965. Circumvention of the mycobactin requirement of Mycobacterium paratuberculosis. Journal of Bacteriology, 89, 762–767.
- NASER, S. A., HULTEN, K., SHAFRAN, I., GRAHAM, D. Y. & EL-ZAATARI, F. A. K., 1999. Specific seroreactivity of Crohn’s patients against P35 and P36 antigens of M. avium subsp. paratuberculosis. Sixth International Colloquium on Paratuberculosis. 14–18 February 1999, Melbourne, Australia.
- NAVARRO, J. A., RAMIS, G., SEVA, J., PALLARES, F. J. & SANCHEZ, J., 1998. Changes in lymphocyte subsets in the intestine and mesenteric lymph nodes on caprine paratuberculosis. Journal of Comparative Pathology, 118, 109–121.
- PAVLIK, I., BEJCKOVA, L., PAVLAS, M., ROZSYPALOVA, Z. & KOSKOVA, S., 1995. Characterization by restriction endonuclease analysis and DNA hybridization using IS900 of bovine, ovine, caprine and human dependent strains of Mycobacterium paratuberculosis isolated in various localities. Veterinary Microbiology, 454, 311-318.
- PAVLIK, I., HORVTHOVA, A., BARTH, J. & RYCHLIK, I., 1996. Study of epidemiology and pathogenesis of paratuberculosis using RFLP (restriction fragment length polymorphism). Proceedings of the Fifth International Colloquium on Paratuberculosis. 29 September to 4 October 1996, Madison, Wisconsin, 202–211.
- PEARSON, J. K. L. & MCCLELLAND, T. G., 1955. Uterine infection and congenital Johne’s disease in cattle. Veterinary Record, 67, 615–616.
- PEMBERTON, D. H., 1979. Diagnosis of Johne’s disease in cattle using mesenteric lymph node biopsy: Accuracy in clinical suspects. Australian Veterinary Journal, 55, 217-219.
- PEREZ, V., GARCIA MARIN, J. F., BRU, R., MORENA, B. & BADIOLA, J. J., 1995. Results of vaccination of adult animals against paratuberculosis. Medicina Veterinaria, 12, 196–201.
- PETRIASAMY, S., TRIPATHI, B. N. & SINGH, N., 2013. Mechanisms of Mycobacterium avium subsp. paratuberculosis induced apoptosis and necrosis in bovine macrophages. Veterinary Microbiology, 165, 392-401.
- PLANTE, Y., REMENDA, B. W., CHELAIK, B. J. & HAINES, D. M., 1996. Detection of Mycobacterium paratuberculosis in formalin-fixed, paraffin-embedded tissues by polymerase chain reaction. Canadian Journal of Veterinary Research, 60, 115-120.
- POUPART, P., COENE, M., VAN HEUVERSWYN, H. & COCITO, C., 1993. Preparation of a specific RNA probe for detection of Mycobacterium paratuberculosis and diagnosis of Johne’s disease. Journal of Clinical Microbiology, 31, 1601–1605.
- RANKIN, J. D., 1962. The experimental infection of cattle with Mycobacterium johnei. IV. Adult cattle maintained in an infectious environment. Journal of Comparative Pathology, 72, 113-117.
- RICHARDS, W. D., 1981. Effects of physical and chemical factors on the viability of, Mycobacterium paratuberculosis. Journal of Clinical Microbiology, 14, 587–588.
- RICHARDS, W. D. & THOEN, C. O., 1977. Effect of freezing on the viability of Mycobacterium paratuberculosis in bovine faeces. Journal of Clinical Microbiology, 6, 392-395.
- RICHARDS, W. D. & WRIGHT, H. S., 1982. Preservation of tissue specimens during transport to mycobacteriology laboratories. Journal of Clinical Microbiology, 17, 393-395.
- RIDEOUT, B. A., BROWN, S. T., DAVIS, W. C., GAY, J. M., GIANNELLA, R. A., HINES, I. M. E., HUESTON, W. D. & HUTCHINSON, L. J., 2003. Control Principles and Programs. Chapter 4. In: The Diagnosis and Control of Johne’s Disease: Committee on the Diagnosis and Control of Johne’s Disease. National Academy of Sciences. National Academy Press, Washington, DC. ISBN: 0-309-08611-6.
- RIDGE, S. E., BADMAN, R. T., MELLOR, A. M. & LARSEN, J. W. A., 1995. Johne’s disease in alpaca (Lama pacos) in Australia. Australian veterinary journal, 72, 150-153.
- RIDGE, S. E. & VIZARD, A. L., 1993. Determination of the optimal cut off value for a serology assay: An example using the Johne’s absorbed ELISA. Journal of Clinical Microbiology, 31, 1256–1261.
- RIS, D. R., HAMEL, K. L. & AYLING, J. M., 1987. Can sheep become infected by grazing pasture contaminated by cattle with Johne’s disease New Zealand Veterinary Journal, 35, 137.
- ROHDE, R. F. & SHULAW, W. P., 1990. Isolation of Mycobacterium paratuberculosis from washed bovine ova after in vitro exposure. American Journal of Veterinary Research, 51, 708–710.
- SALGADO, M., MONTI, G., SEVILLA, I. & MANNING, E., 2014. Association between cattle herd Mycobacterium avium subspecies paratuberculosis (MAP) infection and infection of a hare population. Tropical Animal Health Production, 46, 1313-1316.
- SAXEGAARD, F., 1990. Experimental infection of calves with an apparently specific goat-pathogenic strain of Mycobacterium paratuberculosis. Journal of Comparative Pathology, 98, 31-53.
- SAXEGAARD, F. & BAESS, I., 1988. Relationship between Mycobacterium avium, Mycobacterium paratuberculosis and woodpigeon mycobacteria. Acta Pathologica, Microbiologica et Immunologica Scandinavia, 96, 37-42.
- SEITZ, S. E., HEIDER, L. E., HUESTON, W. D., BECH-NIELSEN, S., RINGS, D. M. & SPANGLER, L., 1989. Bovine fetal infection with Mycobacterium paratuberculosis. Journal of the American Veterinary Medical Association, 194, 1423–1426.
- SELBY, W., 1999. Pathogenesis and therapeutic aspects of Crohn’s disease. Proceedings of the Sixth International Colloquium on Paratuberculosis. 14–18 February 1999. Melbourne, Australia, 515–519.
- SHERMAN, D. M., 1985. Current concepts in Johne’s disease. Veterinary Medicine Small Animal Clinician, 80, 77-84.
- SHERMAN, D. M., MARKHAM, R. J. F. & BATES, F., 1984. Agar-gel immunodiffusion test for the diagnosis of clinical paratuberculosis in cattle. Journal of the American Veterinary Medical Association, 185, 179-182.
- SOCKETT, D. C., CONRAD, T. A., THOMAS, C. B. & COLLINS, M. T., 1992. Evaluation of four serological tests for bovine paratuberculosis. Journal of Clinical Microbiology, 30, 1134–1139.
- STABEL, J. R., 1997. Heat inactivation of Mycobacterium paratuberculosis in raw milk: are current pasteurization conditions effective. Applied and Environmental Microbiology, 63, 4975–4977.
- STABEL, J. R., 1997. An improved method for cultivation of Mycobacterium paratuberculosis from bovine fecal samples and comparison to three other methods. Journal of Veterinary Diagnostic Investigation, 9, 375–380.
- STABEL, J. R., ACKERMANN, M. R. & GOFF, J. P., 1996. Comparison of polyclonal antibodies to three different preparations of Mycobacterium paratuberculosis in immunohistochemical diagnosis of Johne’s disease in cattle. Journal of Veterinary Diagnostic Investigation, 8, 469–473.
- STEVENSON, K., 2015. Genetic diversity of Mycobacterium avium subspecies paratuberculosis and the influnecne of strain type on infection and pathogenesis: a review. Veterinary Research, 46, 64.
- STRINGER, L. A., WILSON, P. R., HEUER, C. & MACKINTOSH, C. G., 2013. A randomised controlled trial of Silirum vaccine for control of paratuberculosis in farmed red deer. doi: 10.1136/vr.101799. The Veterinary Record, 173, 551.
- SUGDEN, E. A., SAMAGH, B. S., CORNER, A. H., TURCOTTE, C., BROOKS, B. W., STEWART, R. B. & DUNCAN, J. R., 1988. Serodiagnosis of ovine paratuberculosis using lipoarabinomannan antigen in an enzyme immuno-assay. Proceedings of the Second International Colloquium on Paratuberculosis. 22 and 23 September, 1988, Maisons-Alfort, 231–235.
- SWEENEY, R. W., JONES, D. E., HABECKER, P. & SCOTT, P., 1998. Interferon-gamma and interleukin 4 gene expression in cows infected with Mycobacterium paratuberculosis. American Journal of Veterinary Research, 59, 842–847.
- SWEENEY, R. W., WHITLOCK, R. H., BOWERSOCK, T. L., CLEARY, D. L., MEINERT, T. R., HABECKER, P. L. & PRUITT, G. W., 2009. Effect of subcutaneous administration of a killed Mycobacterium avium subsp. paratuberculosis vaccine on colonization of tissues following oral exposure to the organism in calves. American Journal of Veterinary Research, 70, 493-497. doi: 10.2460/ajvr.70.4.493.
- SWEENEY, R. W., WHITLOCK, R. H., BUCKLEY, C. L. & SPENCER, P. A., 1995. Evaluation of a commercial enzyme-linked immunosorbent assay for the diagnosis of paratuberculosis in dairy cattle. American Journal of Veterinary Diagnostic Investigation, 7, 488–493.
- SWEENEY, R. W., WHITLOCK, R. H. & ROSENBERGER, A. E., 1992. Mycobacterium paratuberculosis cultured from milk and supramammary lymph nodes of infected asymptomatic cows. Journal of Clinical Microbiology, 30, 166–171.
- SWEENEY, R. W., WHITLOCK, R. H. & ROSENBERGER, A. E., 1992. Mycobacterium paratuberculosis isolated from fetuses of infected cows not manifesting signs of the disease. American Journal of Veterinary Research, 53, 477–480.
- THOEN, C. O. & BAUM, K., 1988. Current knowledge on paratuberculosis. Journal of the American Veterinary Medical Association, 192, 1609–1611.
- THOEN, C. O. & HAAGSMA, J., 1996. Molecular techniques in the diagnosis and control of paratuberculosis in cattle. Journal of the American Veterinary Association, 209, 734–737.
- THOMAS, G. W., 1983. Paratuberculosis in a large goat herd. The Veterinary Record, 113, 464–466.
- THOREL, M. F. & DESMETTRE, P., 1982. Comparative study of mycobactin-dependent strains of mycobacteria, wood-pigeon with Mycobacterium paratuberculosis and Mycobacterium avium. Annales de Microbiologie, 133B, 151–152.
- VAN DER GIESSEN, J. W. B., EGER, A., HAAGSMA, J., HARING, R. M., GAASTRA, W. & VAN DER ZEIJST, B. A. M., 1992. Amplification of 16S rRNA sequences to detect Mycobacterium paratuberculosis. Journal of Medical Microbiology, 36, 255–263.
- VARY, P. W., ANDERSEN, P. R. & GREEN, E., 1990. Use of highly specific DNA probes and the polymerase chain reaction to detect Mycobacterium paratuberculosis in Johne’s disease. Journal of Clinical Microbiology, 28, 932–937.
- VIDAL, S. M., MALO, D., VOGAN, K., SKAMENE, E. & GROSS, P., 1993. Natural resistance to infection with intracellular parasites: Isolation of a candidate for Bcg. Cell, 73, 469–485.
- WADDELL, L., RAJIC, A., STAERK, K. & MCEWEN, S. A., 2016. Mycobacterium avium ssp. paratuberculosis detection in animals, food, water and other sources or vehicles of human exposure: A scoping review of eisting evidence. Preventive Veterinary Medicine, 132, 32-48.
- WHIPPLE, D. L., KAPKE, P. A. & ANDERSEN, P. R., 1992. Comparison of a commercial DNA probe test and three cultivation procedures for detection of Mycobacterium paratuberculosis in bovine faeces. Journal of Veterinary Diagnostic Investigation, 4, 23–27.
- WHIPPLE, D. L., LEFEVURE, R. B., ANDREWS, J. R. I. & THIERMANN, A. B., 1983. Isolation and analysis of restriction endonuclease digestive patterns of chromosomal DNA from Mycobacterium paratuberculosis and other mycobacteria species. Journal of Clinical Microbiology, 25, 1511–1515.
- WHITLOCK, R. H. & BUERGELT, C. D., 1996. Pre-clinical and clinical manifestations of paratuberculosis (including pathology). Veterinary Clinics of North America: Food Animal Practice, 12, 345–355.
- WHITLOCK, R. H., HUTCHISON, L. J., SWEENEY, R. W. & VAN BOSKIRK, M. A., 1991. The Pennsylvania Johne’s control program. Proceedings of the Third International Colloquium on Paratuberculosis. 28 September to 2 October, 1991, Orlando, Florida, 455–465.
- WHITTINGTON, R., WHITTINGTON, A. M., WALDRON, A., BEGG, D. J., DE SILVA, K., PURDIE, A. C. & PLAIN, K. M., 2013. Development and Validation of a Liquid Medium (M7H9C) for Routine Culture of Mycobacterium avium subsp. paratuberculosis to Replace Modified Bactec 12 B Medium. Journal of Clinical Microbiology, 51, 3993-4000.
- WILKS, C. R., TAYLOR, T. K., RUSSEL, E. G. & THURSTON, T. R., 1981. Isolation of mycobacteria induced cross-reactions in the complement fixation test for Johne’s disease. Research in Veterinary Science, 30, 323–327.
- WILLIAMS, E. S., SNYDER, S. P. & MARTIN, K. L., 1983. Pathology of spontaneous and experimental infection of North American wild ruminants with Mycobacterium paratuberculosis. Veterinary Pathology, 20, 274–291.
- WINDSOR, P. A., 2015. Paratuberculosis in sheep and goats. Veterinary Microbiology, 181, 161-169.
- WOOD, P. R., MILNER, A. R., KOPIDAS, K., MACK, W. N. & COATES, K., 1988. Gamma interferon and ELISA titres in cattle infected with Mycobacterium paratuberculosis. Proceedings of the Second International Colloquium on Paratuberculosis.22 and 23 September, 1988, Maisons-Alfort, France, 215–219.
- YOKOMIZO, Y., YUGI, H. & MERKAL, R. S., 1985. A method for avoiding false-positive reactions in an enzyme-linked immunosorbent assay (ELISA) for the diagnosis of bovine paratuberculosis. Japanese Journal of Veterinary Sciences, 47, 111–119.
- ZURBRICK, B. G. & CZUPRYNSKI, C. J., 1987. Ingestion and intracellular growth of Mycobacterium paratuberculosis within blood monocytes and monocyte-derived macrophages. Infection and Immunity, 55, 1588– 1593.




